


Figure
1.
Locations of the sampling stations in the East China Sea.
| Citation: | Linbin Zhou, Yehui Tan, Liangmin Huang. Coral reef ecological pump for gathering and retaining nutrients and exporting carbon: a review and perspectives[J]. Acta Oceanologica Sinica, 2023, 42(6): 1-15. doi: 10.1007/s13131-022-2130-1

|
The East China Sea (ECS) shelf is one of the most extensive continental shelves in the world and most of the ECS is occupied by a continental shelf with depth shallower than 200 m (Yanao and Matsuno, 2013). A number of intricate water masses were found to be the most appropriate choices for studying in the ECS. In the northern part of the ECS, the Changjiang River, which is the prominent source of freshwater, flows southward in spring but northeastward in summer, with a mean outflow of 3×104 m3/s (Yang et al., 2005). The Changjiang River water mixes with seawater to form the Changjiang Diluted Water (CDW), which has the properties of lower temperature and salinity compared to seawater. In addition, as the Changjiang River discharge ranked fourth worldwide, the CDW is the primary source of freshwater mass on this shallow continental shelf (Chen, 2009). The Yellow Sea Coastal Current (YSCC), which influences the northern ECS, flows southward year-round (Wang et al., 2016; Li et al., 2019). In the southern part of the ECS, the Kuroshio water (KW) is the most important element of the ECS circulation system, it’s mainstream flows northeastward along the Okinawa Trough and rejoins the adjacent Pacific Ocean through the Tokara Strait (Lie and Cho, 2016). The KW is a unique carrier of heat, salt and oceanic materials of subtropical open-ocean origin to the ECS (Iseki et al., 2003; Liu et al., 2007; Lie and Cho, 2016).
The formation of water masses throughout the whole ECS region depends on the relative contribution of different factors and processes, such as, inflow of river discharge, Kuroshio, air-sea interaction, sea surface heat exchange, wind stress, and others. In order to better understand the complex processes of water mixing and formation, new insights are needed regarding detailed distributions and characteristics of the water masses of the continental shelf in the ECS.
Water mass analysis and identification have been one of the primary research topics in physical oceanography. After the T-S diagram was introduced by Helland-Hansen (1916), the relationship between temperature and salinity had been applied widely to the study of mixing processes in the ocean (Jacobsen, 1927). Many studies of water masses distribution have used this classical and common method in the ECS (Jacobsen, 1927; Han et al., 2001). Based on the classical T-S diagram, Tomczak (1981) subsequently devised the optimum multiparameter (OMP) method, an inverse modeling technique based on the theory of matrix, for the analysis of distributions of water masses. The OMP method allows for the use of a set of linear mixing equations with hydrological properties (number: m) as the parameters to quantify the mixture ratios of source water masses (number: n), where n>m. It has been successfully applied to oceanic water (Maamaatuaiahutapu et al., 1992, 1994; Klein and Tomczak, 1994; Budillon et al., 2003; Tomczak and Liefrink, 2005), and subsequently become a standard tool for the quantitative description of water mass structures (Dinauer and Mucci, 2018). Besides temperature and salinity, other conservative parameters can also be applied to the OMP method. However, there are two restrictions of the additional parameters: (1) conservative, they must maintain their value in all processes except mixing; (2) independent, any parameter possesses a nonlinear relationship with another parameters (Tomczak, 1981; Thompson and Edwards, 1981; Mackas et al., 1987). Nutrient concentrations, dissolved oxygen, trace elements (e.g., Ba), and radioactive isotopes (e.g., 134Cs, 137Cs and 226Ra) have been used as useful parameters to determine the mixing ratio of water masses (Poole and Tomczak, 1999; Zhang et al., 2007; Gasparin et al., 2014; Liu et al., 2017; Zhao et al., 2018). Whereas, nutrient concentration and dissolved oxygen are not strictly conservation because their distribution are not only subjected to change through circulation and mixing processes, but also affected by the oxidation of biological detritus (Poole and Tomczak, 1999). Furthermore, Chen et al. (1995) pointed out that both temperature and salinity are not strictly conservative in the subtropical marginal seas where precipitation and evaporation vary greatly.
Natural uranium (U) is composed of three alpha radioactive isotopes and their natural abundances are 99.2745% of 238U, 0.7200% of 235U, and 0.0054% of 234U, respectively. The grand-daughter 234U (the half-life period is 245 ka) belongs to the 238U (the half-life period is 4.5 Ga) decay series and is particularly noteworthy, because 234U has an obviously shorter half-life compared to its grand-parent nuclide. A state of secular equilibrium can be achieved in close system after 5 half-lives of 234U. However, there is a common phenomenon discovered since the early 20th century. The 234U and 238U radionuclides in water environment are not in the state of secular equilibrium in the earth’s lithosphere and hydrosphere. This disequilibrium between 234U and 238U in nature was thought to reflect preferential releasing of 234U with respect to its 238U parent into the hydrological environment (Kigoshi, 1971; Sui et al., 2014). The disequilibrium is based on the combined effects, including (1) direct recoil of 234Th (subsequent decay into its daughter 234U) from the mineral grain boundaries, (2) preferential leaching of 234U embedded in recoil tracks (Fleischer, 1980; Andersen et al., 2009) and (3) preferential oxidation of 234U compared to 238U (Adloff and Roessler, 1991). Therefore, terrestrial waters are characterized by the 234U loss from the solid and normally display a representative 234U excess feature. For instance, the modern seawater yields a 234U/238U activity ratio of 1.147 ± 0.001 (Robinson et al., 2004; Andersen et al., 2009), and the river water generally reveals larger 234U excesses with the 234U/238U activity ratio ranging mostly between 1.2 and 1.3 (Chabaux et al., 2003).
Previous studies indicated that the turnover time of the ECS shelf water was approximately 1.3 a, substantially lower than the residence time of U (200–400 ka) in the ocean (Tan et al., 2018; Wang et al., 2018). The 234U/238U activity ratio is different between the oceans and rivers (Andersen et al., 2009). Studies have shown that dissolved U behaves differently in estuaries. For example, results showed conservative behaviors of U in Mississippi River estuaries and Galveston Bay in Texas (Swarzenski and McKee, 1998; Guo et al., 2007). U removal was reported for the Amazon River Estuary (McKee et al., 1987; Swarzenski et al., 1995), the Ganges-Brahmaputra mixing zone (Carroll and Moore, 1993), the southeastern United States (Moore and Shaw, 2008) and the Kalix River Estuary (Porcelli et al., 1997, 2001). Zhou et al. (2016) also reported removal of dissolved U in the Changjiang River Estuary at high salinity. It was also discovered that although the behaviors of dissolved U in estuaries were not uniform, the 234U/238U activity ratio exhibits conservative behavior at high salinity during the mixing process in estuaries and coastal environment (Jiang et al., 2007; Zhou et al., 2016; Liu et al., 2018). The ECS is a river-dominated marginal sea and located between the Asian continent and the Pacific Ocean. Water masses of the ECS bear the signal produced from continent and ocean (Wang et al., 2019). Thus, the 234U/238U activity ratio can be utilized as an additional parameter to identify the contributions of water masses in the ECS. Over recent years, the sensitivity and precision of U isotopic measurements by multi-collector inductively coupled plasma mass spectrometry (MC-ICP-MS) have been further improved. The 234U/238U activity ratio can thus be measured more accurately (Andersen et al., 2009). In this study, we reported three transect profiles of U concentrations and 234U/238U activity ratios. In order to better understand the distribution of the main water masses on the continental shelf of the northern ECS, the relative contributions of water masses were quantified by using salinity, temperature and the 234U/238U activity ratio as the parameters of OMP method.
A total of 96 seawater samples, with 31 samples in vertical profiles, were collected by a CTD rosette system on the R/V Dongfanghong II from March 27 to April 15, 2017. The locations of the sampling stations are shown in Fig. 1. The water depth gradually deepens from west to east. Temperature and salinity of all seawater samples were measured in situ by the Sea-Bird 911 CTD (conductivity-temperature-depth sensor) system instrument.
The seawater samples collected using Niksin bottles were filtered through 0.2 μm membrane (cellulose acetate), and then immediately acidified with concentrated nitric acid (AR grade, Sigma-Aldrich) to pH of less than 2 on board. Finally, the samples were stored in polyethylene plastic bottles under room temperature before laboratory analyses. Chemical pretreatments were performed in a class 1 000 clean lab with class 100 workstations at the Ocean University of China. All samples were spiked with a 233U-236U tracer (Chen et al., 1986; Cheng et al., 2000; Shen et al., 2002). The detailed chemical pretreatments are described in Wang et al. (2017). The recovery of the procedure was better than 90%. The U concentration and isotopic ratio (234U/238U) were analyzed on a MC-ICP-MS (Thermo Fisher Neptune Plus) at the Institute of Geology and Geophysics, Chinese Academy of Sciences. The total procedural blank of 238U was (5±5) pg, which contributed less than 2‰ to the content of U in the measured samples. Precision of 234U/238U with MC-ICP-MS analyses was evaluated with NBS-CRM-112A and was consistently better than 2‰.
The OMP analysis (Tomczak, 1981; Thompson and Edwards, 1981; Mackas et al., 1987; Tomczak and Large, 1989) can be used to determine the mixture ratio of water masses through the parameters of hydrological properties in a certain region, whether the mixing processes are diapycnal or isopycnal mixing. The OMP solves a system of linear equations with the contribution of each source water type (SWT, specific properties characterizing a water mass) as variables and the hydrological properties as the parameters:
| $$ \left[ {\begin{array}{*{20}{c}} 1&1& \cdots &1\\ {{Y_{11}}}&{{Y_{12}}}& \cdots &{{Y_{1n}}}\\ {{Y_{21}}}&{{Y_{22}}}& \cdots &{{Y_{2n}}}\\ \vdots & \vdots & \qquad & \vdots \\ {{Y_{{{m}}1}}}&{{Y_{m2}}}& \cdots &{{Y_{mn}}} \end{array}} \right]\left[ {\begin{array}{*{20}{c}} {{X_1}}\\ {{X_2}}\\ \vdots \\ {{X_n}} \end{array}} \right] = \left[ {\begin{array}{*{20}{c}} 1\\ {Y^{{\rm{obs}}}_1}\\ {Y^{{\rm{obs}}}_2}\\ \vdots \\ {Y^{{\rm{obs}}}_m} \end{array}} \right] + \left[ {\begin{array}{*{20}{c}} {{R_{\rm{M}}}}\\ {{R_{{Y_1}}}}\\ {{R_{{Y_2}}}}\\ \vdots \\ {{R_{{Y_m}}}} \end{array}} \right], $$ | (1) |
where Xi (i = 1, 2, ···, n) is the percentage of contribution of each SWT, Yji (j = 1, 2, ···, m) represents the ith parameter value of the jth SWT, R is residual, and obs refer to the data of observed properties.
The system of the equations can be written in a matrix form:
| $$ {{GX}} = {{d}} + {{R}}, $$ | (2) |
where G is the SWT definition matrix, d is the observation vector, X is the unknown solution vector (ratio of each SWT), and R is the residual vector.
The OMP analysis is constrained to fulfill two physically realistic rules: (1) the mass conservation equation has to be satisfied at any station and (2) the contribution of each SWT has to be positive (Pardo et al., 2012). The problem is solved using a non-negative least squares method. Variations in analytical accuracy and the arbitrariness in SWT parameter choices have to be taken into consideration. Therefore, in order to search for the vector X which minimizes the sum of squares of deviation between measured data and estimates of model parameter (Maamaatuaiahutapu et al., 1992), each equation is weighted by the standard deviation ω of each property (Gasparin et al., 2014). So that Eq. (2) becomes
| $$ {{{R}}^{\rm{T}}}{{R}} = {\left( {{{GX}} - {{d}}} \right)^{\rm{T}}}{{{W}}^{{\bf{ - 1}}}}\left( {{{GX}} - {{d}}} \right){\rm{,}} $$ | (3) |
where T is the transposed matrix operator,
The results of temperature, salinity and U isotopes of the samples from the selected stations are shown in Fig. 2.

The distributions of temperature and salinity in Transects D, F and P are showed in Figs 2a and b, e and f, i and j. There was almost no change in temperature and salinity with depth at all stations of Transect D (Figs 2a and b). The shallower water depth (<100 m) and strong vertical mixing of water masses in the Transect D may account for this unstratified phenomenon (Wei et al., 2013). Due to the influence of the CDW, which receives the fresh water from the Changjiang River, a relatively low salinity (25.81) water sample in the inner shelf was observed at the surface around Station D1. But the extent of the CDW impact was restricted because of the lower Changjiang River discharge during the dry season. At Station D5 (Figs 2a and b), both temperature and salinity were relatively low than those at Stations D3 and D7, which may be related to the intrusion of the YSCC. Wei et al. (2013) analyzed the chemical hydrography at 32.3°N transect between the Yellow Sea and the East China Sea, and testified that the cold deep water in the area of 123.5°–125.5°E originate from YSCC in spring. The high salinity area was observed in the vicinity of 123°E (Fig. 2b), which might be influenced by the KW with high salinity. Zhang et al. (2007) have shown that the KW accounted for a large fraction outside the Changjiang River Estuary.
The temperature and salinity tended to increase from the offshore area toward the open sea in Transect P. In offshore area, the salinity in Transect P increased from Station P1 to Station P7, which was similar to that in Transect D. Compared the salinity vertical profiles of Station P1 (Fig. 2j) with Station D1 (Fig. 2b), a short freshwater plume was found at Station P1. Because Station P1 was deeper and farther away from the estuary than Station D1. The distribution of salinity in the Transect F was different from that of Transects D and P. All salinity values of Transect F are greater than 32.9 (Fig. 2f). The temperature at Station F6 could reach above 20°C (Fig. 2e) because Station F6 was situated in the mainstream and influenced by the KW water with high temperature and salinity.
The vertical profiles of 238U concentrations exhibited extremely similar to the salinity cross-sections, consistent with its conservative feature in the ocean (Ku et al., 1977). The correlation diagram between U concentration and salinity in this study was showed at Fig. 3. The concentration of 238U showed an increase in a linear trend with salinity, suggesting that the U concentration was conservative at definite salinity ranged from 25 to 35 (Fig. 3). Seawater of low 238U concentration presented at Station D1 (Fig. 2c). The 238U concentrations from Transect D range from 2.57 μg/L to 3.14 μg/L, and the two values correspond exactly to the minimum (25.81) and maximum (34.09) salinity. Therefore, the lower concentration of 238U at Station D1 may be caused by the lower value of the riverine U input. The 238U concentration range was 2.78–3.18 μg/L for Transect P and 3.03–3.21 μg/L for Transect F (Fig. 2g). The average 238U concentrations were 3.02 μg/L, 3.11 μg/L and 3.14 μg/L for Transects D, P and F, respectively. The 238U concentrations of Transect D and Transect P were slightly lower than those from Transect F on average. There was a maximum concentration (3.21 μg/L) at intermediate depth (50 m) at Station F5 (Fig. 2g).

The results of 234U/238U activity ratio in the three transects are given in Figs 2d, h, l. As illustrated in Fig. 2d, the activity ratio of the upper-layer water of Station D1 had highest value (1.170 ± 0.002) compared to other samples. Due to the far longer oceanic residence time of U (about 4×105 a) than the ocean circulation time (about 1 000 a), seawater had a quite stable 234U/238U activity ratio of 1.147 ± 0.001 (Dunk et al., 2002). However, Zhou et al. (2015) reported that the range of the average value of 234U/238U activity ratio in the Changjiang River was between 1.374 and 1.424, much higher than the average seawater value. This indicated that the Changjiang River influenced not only the surface water at Station D1, but also the bottom water with a value of 1.159±0.002. The average activity ratio of Station D3 (1.148±0.002) was the same as that of Station D7 (1.148±0.002) (Fig. 2d), and salinity of the two stations was greater than 33, indicating that the waters in the two stations may originate from the same water mass of open ocean. As shown in Fig. 2l, due to the slight influence of the Changjiang River, the 234U/238U activity ratio tended to decrease from the offshore area to the open sea in transect P and eventually reached a stable value of open ocean. The values of 234U/238U activity ratio had relatively small range (1.144–1.151) with an average of 1.147±0.002 in Transect F (Fig. 2h).
In the natural environment, in contrast to nutrients and DO, the fractionation between 234U and 238U is not affected by biogeochemical processes in marine environments, the only way that can cause the fractionation is physical weathering of rock and sediment, which does not occur in the water column. Therefore the 234U/238U activity ratio is a conservation property during water mass mixing, and can be used as an effective parameter in the OMP method for the analysis of water masses, especially when combined with salinity and temperature.
One can modify the above matrix (Eq. (1)) to get a set of new formulas:
| $$ \begin{split} & {X}_{1}+{X}_{2}+{X}_{3}=1+{R}_{\rm{M}}{,} \\ & {X}_{1}{ T}_{1}+{X}_{2}{T}_{2}+{X}_{3}{T}_{3}={T}_{\rm{obs}}+{R}_{{T}}{,} \\ & {X}_{1}{S}_{1}+{X}_{2}{S}_{2}+{X}_{3}{S}_{3}={S}_{\rm{obs}}+{R}_{{S}}{,} \\ & \frac{{\left[{}^{234}{\rm{U}}\right]}_{1}\times {X}_{1}+{\left[{}^{234}{\rm{U}}\right]}_{2}\times {X}_{2}+{\left[{}^{234}{\rm{U}}\right]}_{3}\times {X}_{3}}{{\left[{}^{238}{\rm{U}}\right]}_{1}\times {X}_{1}+{\left[{}^{238}{\rm{U}}\right]}_{2}\times {X}_{2}+{\left[{}^{238}{\rm{U}}\right]}_{3}\times {X}_{3}}{=AR}_{\rm{obs}}{,} \end{split} $$ | (4) |
where [234U] and [238U] are the radioactivities of 234U and 238U, respectively. AR is the activity ratio of [234U] and [238U]. T and S represent the parameters of temperature and salinity, respectively. Other variables are the same as those in Eq. (1).
A water mass is a collection of water parcels defined by a common formation history and has its origin in a specific formation region of the ocean (Tomczak, 1999). Various water masses are able to mix throughout the ocean at different depths so that a given volume of ocean water can be composed of several water masses (Poole and Tomczak, 1999). The OMP analysis requires representation of water masses by SWTs, and the physical and chemical properties of these SWTs were known (Tomczak and Large, 1989; Poole and Tomczak, 1999). A water mass can be represented by a combination of infinite number of SWTs, while it can also be represented by a single SWT in the formation region, as a result of a localized formation process forming the water mass (Tomczak, 1999). Here, we defined the source water mass as a single SWT in the study region. According to recent studies (Qi et al., 2014; Zhou et al., 2018) and the discrimination of water masses in the T-S diagram (Fig. 4), the northern ECS circulation currents are directly governed by the CEW, KW and YSCC.

The water of Station D1 has the typical characteristics of the lowest salinity (25.81), lower temperature (11.37°C) and the highest 234U/238U activity ratio (1.170 7±0.002 0) in all Transect D samples. Thus, Station D1 was used to represent the hydrological variable of the CEW directly. The YSCC traditionally has a temperature of less than 10°C (Ho et al., 1959). Station C2 located in the offshore area of the Yellow Sea, which has lower temperature (9.64°C) and moderate salinity (32.15). So Station C2 was used to represent the YSCC. Three stations (F6, P7 and E7) had a maximum depth of more than 1 000 m in this study and are located in the main stream of the Kuroshio, so they could represent the KW. The red curve in Fig. 4b showed the Kuroshio parameter based on Zhang et al. (2018). The data of Station E7 is consistent with this curve and Station E7 is closer to the upper stream of Kuroshio compared with Stations F6 and P7. Hence, the water at Station E7 is used as KW in this study. The final matrix of SWTs and their characteristic hydrological definitions which were used for input into the OMP method are listed in Table 1.
| Water masses | ω | |||
| CEW | KW | YSCC | ||
| Salinity | 25.81 | 34.81 | 32.15 | 3.5 |
| Temperature/°C | 11.37 | 24.47 | 9.64 | 1.9 |
| 234U/238U activity ratio | 1.170 7 | 1.148 1 | 1.158 4 | 0.005 0 |
| Note: ω is the standard deviation of each property. CEW: estuarine water of Changjiang River, KW: Kuroshio water, YSCC: Yellow Sea Coastal Current. | ||||
 DownLoad:
CSV
DownLoad:
CSV
The vertical distribution of the percentages of water masses contributions calculated in the three transects are shown in Fig. 5. The percentages of CEW to the ECS gradually decrease with the distance away from the estuary, and reached about less than 10% at 123.5°E (Figs 5a, g). This distinct feature is consistent with previous hydrographic observation (Lian et al., 2016; Zhou et al., 2018). The CEW can only affect shallow offshore waters less than 30 m (Fig. 5g). For the entire Transect F, the CEW can be neglected because its contribution was less than 5% of the water fraction of almost all stations (Fig. 5d). These phenomena confirmed that the region affected by the CEW was restricted in the dry season. As illustrated in Fig. 5b, the percentage of YSCC was more than 75% in Transect D except the two stations near the Changjiang River Estuary (Stations D1 and D3). This indicated the YSCC is the dominated water mass in Transect D. In transect P, the high percentage (>65%) of YSCC appeared at the bottom of Stations P3 and P4 (Fig. 5h), indicating that the colder YSCC sinks to the bottom while flows southward in the ECS. Su et al. (1989) have shown that the YSCC has a tendency to sink to the southeast along the continental shelf at the bottom layer. In Transect F, the YSCC decreased gradually from Station F3 to Station F6, and the KW behaved oppositely (Figs 5f, e). Combined with the extremely low percentage of CEW along Transect F (Fig. 5d), the water masses of the Transect F were mainly composed of KW and YSCC. The KW dominated only in the upper water column of Stations F5, F6 and P7 (Figs 5f, i) means the most northern part of ECS was not affected by the KW except the east edge.

Mass conservation residuals were commonly used to evaluate the uncertainty of the OMP analysis. Traditionally, a mass conservation residual (i.e., R) of less than 7% is the standard used to measure the uncertainty of results in OMP practice (Gasparin et al., 2014; Zhou et al., 2018). As shown in Fig. 6, only one datum of R is about 9%, that means more than 97% of the conservation residuals were lower than 6% in this study. Thus, the results of the OMP analysis were reliable (Zhou et al., 2018).
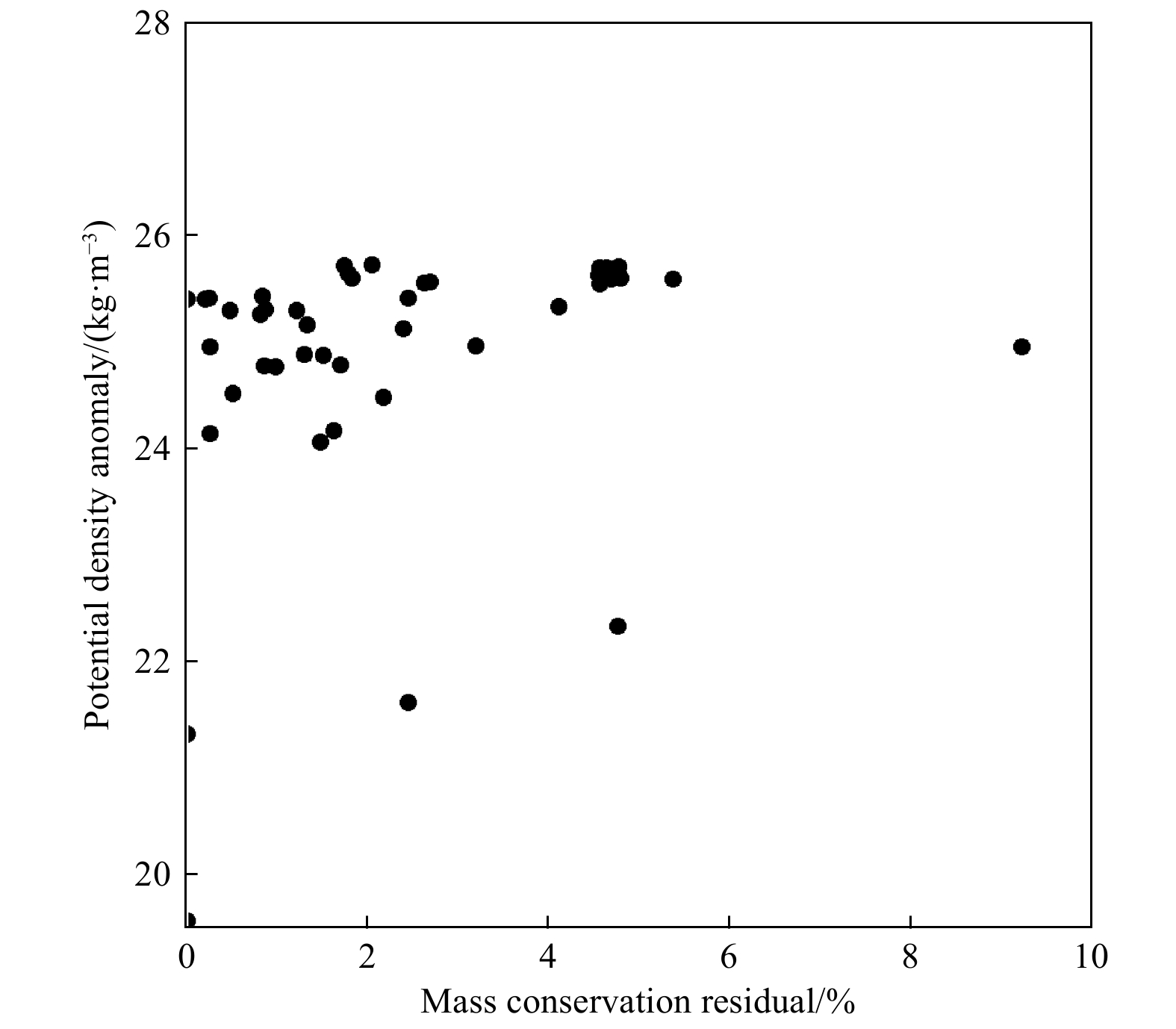
It should be noted that the submarine groundwater discharge (SGD) is also an important water mass contribution to this area (Gu et al., 2012), which was not envolved as a separate end member of this study, because the water in Station D1, which was chosen as the end member of CEW, is actually a mixing water of CDW, SGD and the coastal sea water. The contributions of SGD and other water masses may be envolved in the future research to give a more accurate description of water mixing in this area. Although this study may only represent a snapshot of the water mixing in the northern part of ECS during spring, the activity ratio of 234U/238U has been successfully used as a parameter in the OMP method in the calculation of the water masses mixing in the northern ECS. This method is suitable to the marginal sea area, especially the estuarine region where the end members of water masses usually have different 234U/238U activity ratios, which is a parameter can not be altered by water column biogeochemical processes occurring in the near shore area.
In this study, 234U/238U activity ratios combined with temperature and salinity suggested that the water mass in the northern ECS is a mixture of CEW, KW and YSCC. The mixing ratios of these three water sources were calculated by the OMP method. The mass conservation residuals in the study area were lower than 6%, indicating that the running results of the OMP analysis were reliable. In spring, the dominant source water in the northern ECS was the YSCC. The CEW can only affect shallow offshore waters less than 30 m. Only the east edge of northern ECS was affected by the KW. This result suggested that the 234U/238U activity ratio as a conservative tracer is suitable to characterize and quantify the various water masses in the ECS, as well as other marginal seas influenced by different water masses with their characteristic 234U/238U activity ratios.
We are grateful to Dr. Teng Cong for the help in using MATLAB. We also thank the laboratory staff for their help during sampling and analysis. We thank the Key Laboratory of Marine Chemistry Theory and Technology of Ministry of Education for the assistance (No. 227). This is MCTL contribution No. 142.
|
Albuquerque T, Loiola M, de Anchieta C C Nunes J, et al. 2015. In situ effects of human disturbances on coral reef-fish assemblage structure: temporary and persisting changes are reflected as a result of intensive tourism. Marine and Freshwater Research, 66(1): 23–32. doi: 10.1071/MF13185
|
|
Alldredge A L, Carlson C A, Carpenter R C. 2013. Sources of organic carbon to coral reef flats. Oceanography, 26(3): 108–113. doi: 10.5670/oceanog.2013.52
|
|
Allgeier J E, Burkepile D E, Layman C A. 2017. Animal pee in the sea: consumer-mediated nutrient dynamics in the world’s changing oceans. Global Change Biology, 23(6): 2166–2178. doi: 10.1111/gcb.13625
|
|
Allgeier J E, Layman C A, Mumby P J, et al. 2014. Consistent nutrient storage and supply mediated by diverse fish communities in coral reef ecosystems. Global Change Biology, 20(8): 2459–2472. doi: 10.1111/gcb.12566
|
|
Allgeier J E, Speare K E, Burkepile D E. 2018. Estimates of fish and coral larvae as nutrient subsidies to coral reef ecosystems. Ecosphere, 9(6): e02216
|
|
Allgeier J E, Valdivia A, Cox C, et al. 2016. Fishing down nutrients on coral reefs. Nature Communications, 7: 12461. doi: 10.1038/ncomms12461
|
|
Andradi-Brown D A, Banaszak A T, Frazer T K, et al. 2020. Editorial: coral reefs in the anthropocene—Reflecting on 20 years of reef conservation UK. Frontiers in Marine Science, 7: 364. doi: 10.3389/fmars.2020.00364
|
|
Apprill A. 2020. The role of symbioses in the adaptation and stress responses of marine organisms. Annual Review of Marine Science, 12: 291–314. doi: 10.1146/annurev-marine-010419-010641
|
|
Atkinson M J. 1987. Rates of phosphate uptake by coral reef flat communities. Limnology and Oceanography, 32(2): 426–435. doi: 10.4319/lo.1987.32.2.0426
|
|
Atkinson M J. 1992. Productivity of Enewetak Atoll reef flats predicted from mass transfer relationships. Continental Shelf Research, 12(7–8): 799–807
|
|
Atkinson M J. 2011. Biogeochemistry of nutrients. In: Dubinsky Z, Stambler N, eds. Coral Reefs: An Ecosystem in Transition. Dordrecht: Springer, 199–206
|
|
Atkinson M J, Falter J L. 2003. Coral reefs. In: Black K D, Shimmield G B, eds. Biogeochemistry of Marine Systems. Oxford: Blackwell Publishing, 40–64
|
|
Azam F, Fenchel T, Field J G, et al. 1983. The ecological role of water-column microbes in the sea. Marine Ecology Progress Series, 10: 257–263. doi: 10.3354/meps010257
|
|
Baquiran J I P, Conaco C. 2018. Sponge-microbe partnerships are stable under eutrophication pressure from mariculture. Marine Pollution Bulletin, 136: 125–134. doi: 10.1016/j.marpolbul.2018.09.011
|
|
Bates N R. 2002. Seasonal variability of the effect of coral reefs on seawater CO2 and air-sea CO2 exchange. Limnology and Oceanography, 47(1): 43–52. doi: 10.4319/lo.2002.47.1.0043
|
|
Benavides M, Houlbrèque F, Camps M, et al. 2016. Diazotrophs: a non-negligible source of nitrogen for the tropical coral Stylophora pistillata. Journal of Experimental Biology, 219(17): 2608–2612
|
|
Benkwitt C E, Wilson S K, Graham N A J. 2020. Biodiversity increases ecosystem functions despite multiple stressors on coral reefs. Nature Ecology & Evolution, 4(7): 919–926
|
|
Berkström C, Eggertsen L, Goodell W, et al. 2020. Thresholds in seascape connectivity: the spatial arrangement of nursery habitats structure fish communities on nearby reefs. Ecography, 43(6): 882–896. doi: 10.1111/ecog.04868
|
|
Biegala I C, Raimbault P. 2008. High abundance of diazotrophic picocyanobacteria (<3 µm) in a Southwest Pacific coral lagoon. Aquatic Microbial Ecology, 51: 45–53. doi: 10.3354/ame01185
|
|
Bonnin L, Robbins W D, Boussarie G, et al. 2019. Repeated long-range migrations of adult males in a common Indo-Pacific reef shark. Coral Reefs, 38(6): 1121–1132. doi: 10.1007/s00338-019-01858-w
|
|
Borsa P, Richer De Forges B, Baudat-Franceschi J. 2018. Keep cruises off remote coral reefs. Nature, 558: 372-372
|
|
Bouillon S, Connolly R M. 2009. Carbon exchange among tropical coastal ecosystems. In: Nagelkerken I, ed. Ecological Connectivity among Tropical Coastal Ecosystems. Dordrecht: Springer, 45–70
|
|
Brandl S J, Tornabene L, Goatley C H R, et al. 2019. Demographic dynamics of the smallest marine vertebrates fuel coral reef ecosystem functioning. Science, 364(6446): 1189–1192. doi: 10.1126/science.aav3384
|
|
Brocke H J, Piltz B, Herz N, et al. 2018. Nitrogen fixation and diversity of benthic cyanobacterial mats on coral reefs in Curaçao. Coral Reefs, 37(3): 861–874. doi: 10.1007/s00338-018-1713-y
|
|
Brocke H J, Wenzhoefer F, de Beer D, et al. 2015. High dissolved organic carbon release by benthic cyanobacterial mats in a Caribbean reef ecosystem. Scientific Reports, 5: 8852. doi: 10.1038/srep08852
|
|
Burkepile D E, Allgeier J E, Shantz A A, et al. 2013. Nutrient supply from fishes facilitates macroalgae and suppresses corals in a Caribbean coral reef ecosystem. Scientific Reports, 3: 1493. doi: 10.1038/srep01493
|
|
Callaghan D P, Nielsen P, Cartwright N, et al. 2006. Atoll lagoon flushing forced by waves. Coastal Engineering, 53: 691–704
|
|
Carballo J L, Cruz-Barraza J A, Vega C, et al. 2019. Sponge diversity in Eastern Tropical Pacific coral reefs: an interoceanic comparison. Scientific Reports, 9(1): 9409. doi: 10.1038/s41598-019-45834-4
|
|
Cardini U, Bednarz V N, van Hoytema N, et al. 2016. Budget of primary production and dinitrogen fixation in a highly seasonal Red Sea coral reef. Ecosystems, 19(5): 771–785. doi: 10.1007/s10021-016-9966-1
|
|
Charpy L, Dufour P, Garcia N. 1997. Particulate organic matter in sixteen Tuamotu atoll lagoons (French Polynesia). Marine Ecology Progress Series, 151: 55–65. doi: 10.3354/meps151055
|
|
Chen Xiaoyan, Yu Kefu, Huang Xueyong, et al. 2019. Atmospheric nitrogen deposition increases the possibility of macroalgal dominance on remote coral reefs. Journal of Geophysical Research: Biogeosciences, 124(5): 1355–1369. doi: 10.1029/2019JG005074
|
|
Costa M B, Macedo E C, Valle-Levinson A, et al. 2017. Wave and tidal flushing in a near-equatorial mesotidal atoll. Coral Reefs, 36(1): 277–291. doi: 10.1007/s00338-016-1525-x
|
|
Crossland C J, Hatcher B G, Smith S V. 1991. Role of coral reefs in global ocean production. Coral Reefs, 10(2): 55–64. doi: 10.1007/BF00571824
|
|
Cuet P, Atkinson M J, Blanchot J, et al. 2011. CNP budgets of a coral-dominated fringing reef at La Réunion, France: coupling of oceanic phosphate and groundwater nitrate. Coral Reefs, 30(1): 45–55
|
|
Darwin C R. 1842. The structure and distribution of coral reefs. Being the first part of the geology of the voyage of the Beagle, under the command of Capt. In: Fitzroy R N, ed. During the Years 1832 to 1836. London: Smith Elder and Co.
|
|
Davis S E, Lirman D, Wozniak J R. 2009. Nitrogen and phosphorus exchange among tropical coastal ecosystems. In: Nagelkerken I, ed. Ecological Connectivity Among Tropical Coastal Ecosystems. Dordrecht: Springer, 9–44
|
|
de Goeij J M, van Oevelen D, Vermeij M J A, et al. 2013. Surviving in a marine desert: the sponge loop retains resources within coral reefs. Science, 342(6154): 108–110. doi: 10.1126/science.1241981
|
|
de Paula Y C, Schiavetti A, Sampaio C L S, et al. 2018. The effects of fish feeding by visitors on reef fish in a Marine Protected Area open to tourism. Biota Neotropica, 18(3): e20170339
|
|
Delesalle B, Buscail R, Carbonne J, et al. 1998. Direct measurements of carbon and carbonate export from a coral reef ecosystem (Moorea Island, French Polynesia). Coral Reefs, 17(2): 121–132. doi: 10.1007/s003380050106
|
|
den Haan J, Visser P M, Ganase A E, et al. 2014. Nitrogen fixation rates in algal turf communities of a degraded versus less degraded coral reef. Coral Reefs, 33(4): 1003–1015. doi: 10.1007/s00338-014-1207-5
|
|
Derville S, Torres L G, Dodémont R, et al. 2019. From land and sea, long-term data reveal persistent humpback whale (Megaptera novaeangliae) breeding habitat in New Caledonia. Aquatic Conservation: Marine and Freshwater Ecosystems, 29(10): 1697–1711. doi: 10.1002/aqc.3127
|
|
Diaz-Pulido G, Barrón C. 2020. CO2 enrichment stimulates dissolved organic carbon release in coral reef macroalgae. Journal of Phycology, 56(4): 1039–1052. doi: 10.1111/jpy.13002
|
|
Doty M S, Oguri M. 1956. The island mass effect. ICES Journal of Marine Science, 22(1): 33–37. doi: 10.1093/icesjms/22.1.33
|
|
Du Jianguo, Xie Meiling, Wang Yuyu, et al. 2020. Connectivity of fish assemblages along the mangrove-seagrass-coral reef continuum in Wenchang, China. Acta Oceanologica Sinica, 39(8): 43–52. doi: 10.1007/s13131-019-1490-7
|
|
Dudgeon S, Kübler J E. 2020. A multistressor model of carbon acquisition regulation for macroalgae in a changing climate. Limnology and Oceanography, 65(10): 2541–2555. doi: 10.1002/lno.11470
|
|
Duran A, Collado-Vides L, Burkepile D E. 2016. Seasonal regulation of herbivory and nutrient effects on macroalgal recruitment and succession in a Florida coral reef. PeerJ, 4: e2643. doi: 10.7717/peerj.2643
|
|
Edgar G J, Stuart-Smith R D. 2014. Systematic global assessment of reef fish communities by the Reef Life Survey program. Scientific Data, 1: 140007. doi: 10.1038/sdata.2014.7
|
|
Elsner J B, Kossin J P, Jagger T H. 2008. The increasing intensity of the strongest tropical cyclones. Nature, 455: 92–95
|
|
Erler D V, Santos I R, Eyre B D. 2014. Inorganic nitrogen transformations within permeable carbonate sands. Continental Shelf Research, 77: 69–80. doi: 10.1016/j.csr.2014.02.002
|
|
Eyre B D, Glud R N, Patten N. 2008. Mass coral spawning: a natural large-scale nutrient addition experiment. Limnology and Oceanography, 53: 997–1013
|
|
Fabricius K E. 2005. Effects of terrestrial runoff on the ecology of corals and coral reefs: review and synthesis. Marine Pollution Bulletin, 50(2): 125–146. doi: 10.1016/j.marpolbul.2004.11.028
|
|
Ferrier-Pagès C, Gattuso J P, Cauwet G, et al. 1998. Release of dissolved organic carbon and nitrogen by the zooxanthellate coral Galaxea fascicularis. Marine Ecology Progress Series, 172: 265–274. doi: 10.3354/meps172265
|
|
Ferrier-Pagès C, Leclercq N, Jaubert J, et al. 2000. Enhancement of pico- and nanoplankton growth by coral exudates. Aquatic Microbial Ecology, 21: 203–209. doi: 10.3354/ame021203
|
|
Ferrier-Pagès C, Sauzéat L, Balter V. 2018. Coral bleaching is linked to the capacity of the animal host to supply essential metals to the symbionts. Global Change Biology, 24: 3145–3157
|
|
Fiore C L, Jarett J K, Olson N D, et al. 2010. Nitrogen fixation and nitrogen transformations in marine symbioses. Trends in Microbiology, 18(10): 455–463. doi: 10.1016/j.tim.2010.07.001
|
|
Fitt W K, McFarland F K, Warner M E, et al. 2000. Seasonal patterns of tissue biomass and densities of symbiotic dinoflagellates in reef corals and relation to coral bleaching. Limnology and Oceanography, 45(3): 677–685. doi: 10.4319/lo.2000.45.3.0677
|
|
Ford A K, Van Hoytema N, Moore B R, et al. 2017. High sedimentary oxygen consumption indicates that sewage input from small islands drives benthic community shifts on overfished reefs. Environmental Conservation, 44(4): 405–411. doi: 10.1017/S0376892917000054
|
|
Fox M D, Elliott Smith E A, Smith J E, et al. 2019. Trophic plasticity in a common reef-building coral: insights from δ13C analysis of essential amino acids. Functional Ecology, 33(11): 2203–2214. doi: 10.1111/1365-2435.13441
|
|
Francis F T, Cȏté I M. 2018. Fish movement drives spatial and temporal patterns of nutrient provisioning on coral reef patches. Ecosphere, 9(5): e02225
|
|
Friedlander A M, Obura D, Aumeeruddy R, et al. 2014. Coexistence of low coral cover and high fish biomass at Farquhar Atoll, Seychelles. PLoS ONE, 9(1): e87359. doi: 10.1371/journal.pone.0087359
|
|
Furnas M J, Mitchell A W, Skuza M. 1995. Nitrogen and Phosphorus Budgets for the Central Great Barrier Reef Shelf. Towmsville Queensland: Great Barrier Reef Marine Park Authority
|
|
Gattuso J P, Frankignoulle M, Smith S V. 1999. Measurement of community metabolism and significance in the coral reef CO2 source-sink debate. Proceedings of the National Academy of Sciences of the United States of America, 96(23): 13017–13022. doi: 10.1073/pnas.96.23.13017
|
|
Geesey G G, Alexander G V, Bray R N, et al. 1984. Fish fecal pellets are a source of minerals for inshore reef communities. Marine Ecology Progress Series, 15: 19–25. doi: 10.3354/meps015019
|
|
Genin A, Monismith S G, Reidenbach M A, et al. 2009. Intense benthic grazing of phytoplankton in a coral reef. Limnology and Oceanography, 54(3): 938–951. doi: 10.4319/lo.2009.54.3.0938
|
|
Goatley C H R, Bellwood D R. 2016. Body size and mortality rates in coral reef fishes: a three-phase relationship. Proceedings of the Royal Society B: Biological Sciences, 283(1841): 20161858. doi: 10.1098/rspb.2016.1858
|
|
Godinot C, Gaysinski M, Thomas O P, et al. 2016. On the use of 31P NMR for the quantification of hydrosoluble phosphorus-containing compounds in coral host tissues and cultured zooxanthellae. Scientific Reports, 6: 21760. doi: 10.1038/srep21760
|
|
González-Dávila M, Casiano J M S, Machín F. 2017. Changes in the partial pressure of carbon dioxide in the Mauritanian-Cap Vert upwelling region between 2005 and 2012. Biogeosciences, 14(17): 3859–3871. doi: 10.5194/bg-14-3859-2017
|
|
Gove J M, McManus M A, Neuheimer A B, et al. 2016. Near-island biological hotspots in barren ocean basins. Nature Communications, 7: 10581. doi: 10.1038/ncomms10581
|
|
Graham N A J, Nash K L. 2013. The importance of structural complexity in coral reef ecosystems. Coral Reefs, 32(2): 315–326. doi: 10.1007/s00338-012-0984-y
|
|
Graham N A J, Wilson S K, Carr P, et al. 2018. Seabirds enhance coral reef productivity and functioning in the absence of invasive rats. Nature, 559(7713): 250–253. doi: 10.1038/s41586-018-0202-3
|
|
Gray S E C, DeGrandpre M D, Langdon C, et al. 2012. Short-term and seasonal pH, pCO2 and saturation state variability in a coral-reef ecosystem. Global Biogeochemical Cycles, 26(3): GB3012
|
|
Green R H, Jones N L, Rayson M D, et al. 2019. Nutrient fluxes into an isolated coral reef atoll by tidally driven internal bores. Limnology and Oceanography, 64(2): 461–473. doi: 10.1002/lno.11051
|
|
Green A L, Maypa A P, Almany G R, et al. 2015. Larval dispersal and movement patterns of coral reef fishes, and implications for marine reserve network design. Biological Reviews, 90(4): 1215–1247. doi: 10.1111/brv.12155
|
|
Grover R, Ferrier-Pagès C, Maguer J F, et al. 2014. Nitrogen fixation in the mucus of Red Sea corals. Journal of Experimental Biology, 217(22): 3962–3963
|
|
Gruber R K, Lowe R J, Falter J L. 2018. Benthic uptake of phytoplankton and ocean-reef exchange of particulate nutrients on a tide-dominated reef. Limnology and Oceanography, 63(4): 1545–1561. doi: 10.1002/lno.10790
|
|
Haas A F, Fairoz M F M, Kelly L W, et al. 2016. Global microbialization of coral reefs. Nature Microbiology, 1: 16042
|
|
Hadaidi G, Gegner H M, Ziegler M, et al. 2019. Carbohydrate composition of mucus from scleractinian corals from the central Red Sea. Coral Reefs, 38(1): 21–27. doi: 10.1007/s00338-018-01758-5
|
|
Hamner W M, Colin P L, Hamner P P. 2007. Export-import dynamics of zooplankton on a coral reef in Palau. Marine Ecology Progress Series, 334: 83–92
|
|
Hamner W M, Jones M S, Carleton J H, et al. 1988. Zooplankton, planktivorous fish, and water currents on a windward reef face: Great Barrier Reef, Australia. Bulletin of Marine Science, 42(3): 459–479
|
|
Hata H, Kudo S, Yamano H, et al. 2002. Organic carbon flux in Shiraho coral reef (Ishigaki Island, Japan). Marine Ecology Progress Series, 232: 129–140. doi: 10.3354/meps232129
|
|
Hata H, Suzuki A, Maruyama T, et al. 1998. Carbon flux by suspended and sinking particles around the barrier reef of Palau, western Pacific. Limnology and Oceanography, 43(8): 1883–1893. doi: 10.4319/lo.1998.43.8.1883
|
|
Hatcher B G. 1988. Coral reef primary productivity: a beggar’s banquet. Trends in Ecology & Evolution, 3(5): 106–111
|
|
Hatcher B G. 1997. Coral reef ecosystems: how much greater is the whole than the sum of the parts?. Coral Reefs, 16: S77–S91
|
|
Hernández-León S. 1991. Accumulation of mesozooplankton in a wake area as a causative mechanism of the “island-mass effect”. Marine Biology, 109(1): 141–147. doi: 10.1007/BF01320241
|
|
Heupel M R, Papastamatiou Y P, Espinoza M, et al. 2019. Reef shark science—key questions and future directions. Frontiers in Marine Science, 6: 12. doi: 10.3389/fmars.2019.00012
|
|
Hoer D R, Gibson P J, Tommerdahl J P, et al. 2018. Consumption of dissolved organic carbon by Caribbean reef sponges. Limnology and Oceanography, 63(1): 337–351. doi: 10.1002/lno.10634
|
|
Honig S E, Mahoney B. 2016. Evidence of seabird guano enrichment on a coral reef in Oahu, Hawaii. Marine Biology, 163(2): 22. doi: 10.1007/s00227-015-2808-4
|
|
Houlbrèque F, Ferrier-Pagès C. 2009. Heterotrophy in tropical scleractinian corals. Biological Reviews, 84(1): 1–17. doi: 10.1111/j.1469-185X.2008.00058.x
|
|
Huang Liangmin. 1991. Characteristics of photosynthetic pigments and primary productivity distribution in the waters around Nansha Islands (In Chinese with English abstract). In: Chen Q, Ed. Mutidisciplinary Expedition to Nansha Islands Chinese Academy of Sciences, eds. Proceedings of Studies on Marine Organisms in Nansha Islands and Adjacent Waters II. Beijing: China Ocean Press, 34–49
|
|
Huang Liangmin. 1997. A Study on Ecological Processes of Nansha Islands Sea Area I (in Chinese). Beijing: Science Press
|
|
Huang Liangmin, Tan Yehui, Song Xingyu, et al. 2020. A Study on Ecological Processes of Nansha Islands Sea Area (in Chinese). Beijing: Science Press
|
|
Ikawa H, Faloona I, Kochendorfer J, et al. 2013. Air-sea exchange of CO2 at a Northern California coastal site along the California Current upwelling system. Biogeosciences, 10(7): 4419–4432. doi: 10.5194/bg-10-4419-2013
|
|
James A K, Washburn L, Gotschalk C, et al. 2020. An island mass effect resolved near Mo’orea, French Polynesia. Frontiers in Marine Science, 7: 16. doi: 10.3389/fmars.2020.00016
|
|
Jantzen C, Wild C, Rasheed M, et al. 2010. Enhanced pore-water nutrient fluxes by the upside-down jellyfish Cassiopea sp. in a Red Sea coral reef. Marine Ecology Progress Series, 411: 117–125. doi: 10.3354/meps08623
|
|
Jiang Wei, Yang Haodan, Wu Xingyuan, et al. 2020. Research progress of environmental influence and coral record of submarine groundwater discharge in coral reefs. Haiyang Xuebao (In Chinese with English Abstract), 42(11): 1–11. doi: 10.3969/j.issn.0253-4193.2020.11.001
|
|
Johannes R E, Alberts J, D’Elia C, et al. 1972. The metabolism of some coral reef communities: a team study of nutrient and energy flux at Eniwetok. BioScience, 22(9): 541–543. doi: 10.2307/1296314
|
|
Johannes R E, Coles S L, Kuenzel N T. 1970. The role of zooplankton in the nutrition of some scleractinian corals. Limnology and Oceanography, 15(4): 579–586. doi: 10.4319/lo.1970.15.4.0579
|
|
Karcher D B, Roth F, Carvalho S, et al. 2020. Nitrogen eutrophication particularly promotes turf algae in coral reefs of the central Red Sea. PeerJ, 8: e8737. doi: 10.7717/peerj.8737
|
|
Karnauskas K B, Cohen A L. 2012. Equatorial refuge amid tropical warming. Nature Climate Change, 2: 530–534
|
|
Kayanne H, Suzuki A, Saito H. 1995. Diurnal changes in the partial pressure of carbon dioxide in coral reef water. Science, 269(5221): 214–216. doi: 10.1126/science.269.5221.214
|
|
Ke Zhixin, Tan Yehui, Huang Liangmin, et al. 2018. Spatial distribution patterns of phytoplankton biomass and primary productivity in six coral atolls in the central South China Sea. Coral Reefs, 37(3): 919–927. doi: 10.1007/s00338-018-1717-7
|
|
Kealoha A K, Shamberger K E F, Reid E C, et al. 2019. Heterotrophy of oceanic particulate organic matter elevates net ecosystem calcification. Geophysical Research Letters, 46(16): 9851–9860. doi: 10.1029/2019GL083726
|
|
Khan J A, Goatley C H R, Brandl S J, et al. 2017. Shelter use by large reef fishes: long-term occupancy and the impacts of disturbance. Coral Reefs, 36(4): 1123–1132. doi: 10.1007/s00338-017-1604-7
|
|
Kinsey D W, Hopley D. 1991. The significance of coral reefs as global carbon sinks—response to Greenhouse. Global and Planetary Change, 3(4): 363–377. doi: 10.1016/0921-8181(91)90117-F
|
|
Kossin J P, Knapp K R, Olander T L, et al. 2020. Global increase in major tropical cyclone exceedance probability over the past four decades. Proceedings of the National Academy of Sciences of the United States of America, 117(22): 11975–11980. doi: 10.1073/pnas.1920849117
|
|
Kötter I. 2003. Feeding ecology of coral reef sponges [dissertation]. Bremen: University Bremen, 77
|
|
Kumagai N H, Molinos J G, Yamano H, et al. 2018. Ocean currents and herbivory drive macroalgae-to-coral community shift under climate warming. Proceedings of the National Academy of Sciences of the United States of America, 115(36): 8990–8995. doi: 10.1073/pnas.1716826115
|
|
Larkum A W D, Kennedy I R, Muller W J. 1988. Nitrogen fixation on a coral reef. Marine Biology, 98(1): 143–155. doi: 10.1007/BF00392669
|
|
Lee S, Ford A K, Mangubhai S, et al. 2018. Effects of sandfish (Holothuria scabra) removal on shallow-water sediments in Fiji. PeerJ, 6: e4773. doi: 10.7717/peerj.4773
|
|
Leichter J J, Stewart H L, Miller S L. 2003. Episodic nutrient transport to Florida coral reefs. Limnology and Oceanography, 48: 1394–1407
|
|
Lesser M P, Morrow K M, Pankey S M, et al. 2018. Diazotroph diversity and nitrogen fixation in the coral Stylophora pistillata from the Great Barrier Reef. The ISME Journal, 12(3): 813–824. doi: 10.1038/s41396-017-0008-6
|
|
Leys S P, Yahel G, Reidenbach M A, et al. 2011. The sponge pump: the role of current induced flow in the design of the sponge body plan. PLoS ONE, 6(12): e27787. doi: 10.1371/journal.pone.0027787
|
|
Liberman T, Genin A, Loya Y. 1995. Effects on growth and reproduction of the coral Stylophora pistillata by the mutualistic damselfish Dascyllus marginatus. Marine Biology, 121(4): 741–746. doi: 10.1007/BF00349310
|
|
Liu Qingxia, Zhou Linbin, Wu Yun, et al. 2022. Quantification of the carbon released by a marine fish using a carbon release model and radiocarbon. Marine Pollution Bulletin, 181: 113908. doi: 10.1016/j.marpolbul.2022.113908
|
|
Lønborg C, Calleja M L, Fabricius K E, et al. 2019. The Great Barrier Reef: a source of CO2 to the atmosphere. Marine Chemistry, 210: 24–33. doi: 10.1016/j.marchem.2019.02.003
|
|
Lønborg C, Doyle J, Furnas M, et al. 2017. Seasonal organic matter dynamics in the Great Barrier Reef lagoon: contribution of carbohydrates and proteins. Continental Shelf Research, 138: 95–105. doi: 10.1016/j.csr.2017.01.010
|
|
Lorrain A, Houlbrèque F, Benzoni F, et al. 2017. Seabirds supply nitrogen to reef-building corals on remote Pacific islets. Scientific Reports, 7(1): 3721. doi: 10.1038/s41598-017-03781-y
|
|
Martínez-Moreno J, Hogg A M, England M H, et al. 2021. Global changes in oceanic mesoscale currents over the satellite altimetry record. Nature Climate Change, 11(5): 397–403. doi: 10.1038/s41558-021-01006-9
|
|
Massaro R F S, De Carlo E H, Drupp P S, et al. 2012. Multiple factors driving variability of CO2 exchange between the ocean and atmosphere in a tropical coral reef environment. Aquatic Geochemistry, 18(4): 357–386. doi: 10.1007/s10498-012-9170-7
|
|
McMahon A, Santos I R. 2017. Nitrogen enrichment and speciation in a coral reef lagoon driven by groundwater inputs of bird guano. Journal of Geophysical Research: Oceans, 122(9): 7218–7236. doi: 10.1002/2017JC012929
|
|
McManus M A, Benoit-Bird K J, Brock Woodson C. 2008. Behavior exceeds physical forcing in the diel horizontal migration of the midwater sound-scattering layer in Hawaiian waters. Marine Ecology Progress Series, 365: 91–101. doi: 10.3354/meps07491
|
|
McMurray S E, Stubler A D, Erwin P M, et al. 2018. A test of the sponge-loop hypothesis for emergent Caribbean reef sponges. Marine Ecology Progress Series, 588: 1–14. doi: 10.3354/meps12466
|
|
Meunier V, Bonnet S, Pernice M, et al. 2019. Bleaching forces coral’s heterotrophy on diazotrophs and Synechococcus. The ISME Journal, 13(11): 2882–2886. doi: 10.1038/s41396-019-0456-2
|
|
Meyer J L, Schultz E T. 1985. Tissue condition and growth rate of corals associated with schooling fish. Limnology and Oceanography, 30(1): 157–166. doi: 10.4319/lo.1985.30.1.0157
|
|
Meyer J L, Schultz E T, Helfman G S. 1983. Fish schools: an asset to corals. Science, 220(4601): 1047–1049. doi: 10.1126/science.220.4601.1047
|
|
Mueller B, den Haan J, Visser P M, et al. 2016. Effect of light and nutrient availability on the release of dissolved organic carbon (DOC) by Caribbean turf algae. Scientific Reports, 6: 23248. doi: 10.1038/srep23248
|
|
Nagelkerken I. 2009. Ecological Connectivity Among Tropical Coastal Ecosystems. Dordrecht: Springer
|
|
Nelson C E, Alldredge A L, McCliment E A, et al. 2011. Depleted dissolved organic carbon and distinct bacterial communities in the water column of a rapid-flushing coral reef ecosystem. The ISME Journal, 5(8): 1374–1387. doi: 10.1038/ismej.2011.12
|
|
Newman M J H, Paredes G A, Sala E, et al. 2006. Structure of Caribbean coral reef communities across a large gradient of fish biomass. Ecology Letters, 9(11): 1216–1227. doi: 10.1111/j.1461-0248.2006.00976.x
|
|
Ning Zhiming, Yu Kefu, Wang Yinghui, et al. 2019. Carbon and nutrient dynamics of permeable carbonate and silicate sands adjacent to coral reefs around Weizhou Island in the northern South China Sea. Estuarine, Coastal and Shelf Science, 225: 106229
|
|
Odum E P, Barrett G W. 2005. Fundamentals of Ecology. 5th ed. Belmont: Thomson Brooks/Cole
|
|
Odum H T, Odum E P. 1955. Trophic structure and productivity of a windward coral reef community on Eniwetok atoll. Ecological Monographs, 25(3): 291–320. doi: 10.2307/1943285
|
|
Palacios D M. 2002. Factors influencing the island-mass effect of the Galápagos Archipelago. Geophysical Research Letters, 29(23): 2134
|
|
Patten N L, Wyatt A S J, Lowe R J, et al. 2011. Uptake of picophytoplankton, bacterioplankton and virioplankton by a fringing coral reef community (Ningaloo Reef, Australia). Coral Reefs, 30(3): 555–567. doi: 10.1007/s00338-011-0777-8
|
|
Pawlik J R, Loh T L, McMurray S E. 2018. A review of bottom-up vs. top-down control of sponges on Caribbean fore-reefs: what’s old, what’s new, and future directions. PeerJ, 6: e4343. doi: 10.7717/peerj.4343
|
|
Pawlik J R, McMurray S E. 2020. The emerging ecological and biogeochemical importance of sponges on coral reefs. Annual Review of Marine Science, 12: 315–337
|
|
Peel L R, Daly R, Daly C A K, et al. 2019. Stable isotope analyses reveal unique trophic role of reef manta rays (Mobula alfredi) at a remote coral reef. Royal Society Open Science, 6(9): 190599. doi: 10.1098/rsos.190599
|
|
Peirano A, Morri C, Bianchi C N, et al. 2001. Biomass, carbonate standing stock and production of the Mediterranean coral Cladocora caespitosa (L. ). Facies, 44(1): 75–80. doi: 10.1007/BF02668168
|
|
Pinnegar J K, Polunin N V C. 2006. Planktivorous damselfish support significant nitrogen and phosphorus fluxes to Mediterranean reefs. Marine Biology, 148(5): 1089–1099. doi: 10.1007/s00227-005-0141-z
|
|
Polovina J J. 1984. Model of a coral reef ecosystem I. The ECOPATH model and its application to French Frigate Shoals. Coral Reefs, 3(1): 1–11. doi: 10.1007/BF00306135
|
|
Pupier C A, Bednarz V N, Grover R, et al. 2019. Divergent capacity of scleractinian and soft corals to assimilate and transfer diazotrophically derived nitrogen to the reef environment. Frontiers in Microbiology, 10: 1860. doi: 10.3389/fmicb.2019.01860
|
|
Purcell S W, Conand C, Uthicke S, et al. 2016. Ecological roles of exploited sea cucumbers. In: Hughes R N, Hughes D J, Smith I P, et al., eds. Oceanography and Marine Biology: An Annual Review, Vol. 54. Boca Raton: CRC Press, 367–386
|
|
Qiu Dajun, Huang Liangmin, Huang Hui, et al. 2010. Two functionally distinct ciliates dwelling in Acropora corals in the South China Sea near Sanya, Hainan Province, China. Applied and Environmental Microbiology, 76(16): 5639–5643. doi: 10.1128/AEM.03009-09
|
|
Rädecker N, Pogoreutz C, Voolstra C R, et al. 2015. Nitrogen cycling in corals: the key to understanding holobiont functioning?. Trends in Microbiology, 23(8): 490–497
|
|
Radice V Z, Hoegh-Guldberg O, Fry B, et al. 2019. Upwelling as the major source of nitrogen for shallow and deep reef-building corals across an oceanic atoll system. Functional Ecology, 33(6): 1120–1134. doi: 10.1111/1365-2435.13314
|
|
Reid S B, Hirota J, Young R E, et al. 1991. Mesopelagic-boundary community in Hawaii: micronekton at the interface between neritic and oceanic ecosystems. Marine Biology, 109(3): 427–440. doi: 10.1007/BF01313508
|
|
Ribes M, Coma R, Atkinson M J, et al. 2005. Sponges and ascidians control removal of particulate organic nitrogen from coral reef water. Limnology and Oceanography, 50(5): 1480–1489. doi: 10.4319/lo.2005.50.5.1480
|
|
Rice M M, Maher R L, Vega Thurber R, et al. 2019. Different nitrogen sources speed recovery from corallivory and uniquely alter the microbiome of a reef-building coral. PeerJ, 7: e8056
|
|
Richardson T L. 2019. Mechanisms and pathways of small-phytoplankton export from the surface ocean. Annual Review of Marine Science, 11: 57–74. doi: 10.1146/annurev-marine-121916-063627
|
|
Richardson T L, Jackson G A. 2007. Small phytoplankton and carbon export from the surface ocean. Science, 315(5813): 838–840. doi: 10.1126/science.1133471
|
|
Richter C, Wunsch M, Rasheed M, et al. 2001. Endoscopic exploration of Red Sea coral reefs reveals dense populations of cavity-dwelling sponges. Nature, 413(6857): 726–730. doi: 10.1038/35099547
|
|
Rix L, de Goeij J M, van Oevelen D, et al. 2017. Differential recycling of coral and algal dissolved organic matter via the sponge loop. Functional Ecology, 31(3): 778–789. doi: 10.1111/1365-2435.12758
|
|
Rix L, de Goeij J M, van Oevelen D, et al. 2018. Reef sponges facilitate the transfer of coral-derived organic matter to their associated fauna via the sponge loop. Marine Ecology Progress Series, 589: 85–96. doi: 10.3354/meps12443
|
|
Roopin M, Henry R P, Chadwick N E. 2008. Nutrient transfer in a marine mutualism: patterns of ammonia excretion by anemonefish and uptake by giant sea anemones. Marine Biology, 154(3): 547–556. doi: 10.1007/s00227-008-0948-5
|
|
Ruiz Sebastián C, McClanahan T R. 2013. Description and validation of production processes in the coral reef ecosystem model CAFFEE (Coral–Algae–Fish-Fisheries Ecosystem Energetics) with a fisheries closure and climatic disturbance. Ecological Modelling, 263: 326–348. doi: 10.1016/j.ecolmodel.2013.05.012
|
|
Sambrook K, Bonin M C, Bradley M, et al. 2020. Broadening our horizons: seascape use by coral reef-associated fishes in Kavieng, Papua New Guinea, is common and diverse. Coral Reefs, 39(4): 1187–1197. doi: 10.1007/s00338-020-01954-2
|
|
Sandin S A, Smith J E, Demartini E E, et al. 2008. Baselines and degradation of coral reefs in the northern Line Islands. PLoS ONE, 3(2): e1548. doi: 10.1371/journal.pone.0001548
|
|
Savage C. 2019. Seabird nutrients are assimilated by corals and enhance coral growth rates. Scientific Reports, 9(1): 4284. doi: 10.1038/s41598-019-41030-6
|
|
Schöttner S, Pfitzner B, Grünke S, et al. 2011. Drivers of bacterial diversity dynamics in permeable carbonate and silicate coral reef sands from the Red Sea. Environmental Microbiology, 13(7): 1815–1826. doi: 10.1111/j.1462-2920.2011.02494.x
|
|
Sebens K P, Vandersall K S, Savina L A, et al. 1996. Zooplankton capture by two scleractinian corals, Madracis mirabilis and Montastrea cavernosa, in a field enclosure. Marine Biology, 127(2): 303–317. doi: 10.1007/BF00942116
|
|
Shantz A A, Ladd M C, Schrack E, et al. 2015. Fish-derived nutrient hotspots shape coral reef benthic communities. Ecological Applications, 25(8): 2142–2152. doi: 10.1890/14-2209.1
|
|
Sheng Huaxia, Wan X S, Zou Bobo, et al. 2023. An efficient diazotroph-derived nitrogen transfer pathway in coral reef system. Limnology and Oceanography, 68: 963–981
|
|
Shiozaki T, Kodama T, Furuya K. 2014. Large-scale impact of the island mass effect through nitrogen fixation in the western South Pacific Ocean. Geophysical Research Letters, 41(8): 2907–2913. doi: 10.1002/2014GL059835
|
|
Silveira C B, Silva-Lima A W, Francini-Filho R B, et al. 2015. Microbial and sponge loops modify fish production in phase-shifting coral reefs. Environmental Microbiology, 17(10): 3832–3846. doi: 10.1111/1462-2920.12851
|
|
Starke C, Ekau W, Moosdorf N. 2020. Enhanced productivity and fish abundance at a submarine spring in a coastal lagoon on Tahiti, French Polynesia. Frontiers in Marine Science, 6: 809. doi: 10.3389/fmars.2019.00809
|
|
Staunton Smith J, Johnson C R. 1995. Nutrient inputs from seabirds and humans on a populated coral cay. Marine Ecology Progress Series, 124: 189–200. doi: 10.3354/meps124189
|
|
Suzuki A, Kawahata H. 2003. Carbon budget of coral reef systems: an overview of observations in fringing reefs, barrier reefs and atolls in the Indo-Pacific regions. Tellus B: Chemical and Physical Meteorology, 55(2): 428–444. doi: 10.3402/tellusb.v55i2.16761
|
|
Suzuki A, Kawahata H, Ayukai T, et al. 2001. The oceanic CO2 system and carbon budget in the Great Barrier Reef, Australia. Geophysical Research Letters, 28(7): 1243–1246. doi: 10.1029/2000GL011875
|
|
Tait D R, Erler D V, Santos I R, et al. 2014. The influence of groundwater inputs and age on nutrient dynamics in a coral reef lagoon. Marine Chemistry, 166: 36–47. doi: 10.1016/j.marchem.2014.08.004
|
|
Tanaka Y, Miyajima T, Koike I, et al. 2008. Production of dissolved and particulate organic matter by the reef-building corals Porites cylindrica and Acropora pulchra. Bulletin of Marine Science, 82(2): 237–245
|
|
Tanaka Y, Miyajima T, Watanabe A, et al. 2011a. Distribution of dissolved organic carbon and nitrogen in a coral reef. Coral Reefs, 30(2): 533–541. doi: 10.1007/s00338-011-0735-5
|
|
Tanaka Y, Ogawa H, Miyajima T. 2011b. Production and bacterial decomposition of dissolved organic matter in a fringing coral reef. Journal of Oceanography, 67(4): 427–437. doi: 10.1007/s10872-011-0046-z
|
|
Thornhill D J, Rotjan R D, Todd B D, et al. 2011. A connection between colony biomass and death in Caribbean reef-building corals. PLoS ONE, 6(12): e29535. doi: 10.1371/journal.pone.0029535
|
|
Tilstra A, van Hoytema N, Cardini U, et al. 2018. Effects of water column mixing and stratification on planktonic primary production and dinitrogen fixation on a northern Red Sea coral reef. Frontiers in Microbiology, 9: 2351. doi: 10.3389/fmicb.2018.02351
|
|
Torréton J P, Dufour P. 1996. Temporal and spatial stability of bacterioplankton biomass and productivity in an atoll lagoon. Aquatic Microbial Ecology, 11: 251–261. doi: 10.3354/ame011251
|
|
Torréton J P, Pagès J, Talbot V. 2002. Relationships between bacterioplankton and phytoplankton biomass, production and turnover rate in Tuamotu atoll lagoons. Aquatic Microbial Ecology, 28: 267–277. doi: 10.3354/ame028267
|
|
Turk-Kubo K A, Frank I E, Hogan M E, et al. 2015. Diazotroph community succession during the VAHINE mesocosm experiment (New Caledonia lagoon). Biogeosciences, 12(24): 7435–7452. doi: 10.5194/bg-12-7435-2015
|
|
Uthicke S. 2001a. Interactions between sediment-feeders and microalgae on coral reefs: grazing losses versus production enhancement. Marine Ecology Progress Series, 210: 125–138. doi: 10.3354/meps210125
|
|
Uthicke S. 2001b. Nutrient regeneration by abundant coral reef holothurians. Journal of Experimental Marine Biology and Ecology, 265(2): 153–170. doi: 10.1016/S0022-0981(01)00329-X
|
|
Vega Thurber R L, Burkepile D E, Fuchs C, et al. 2014. Chronic nutrient enrichment increases prevalence and severity of coral disease and bleaching. Global Change Biology, 20: 544–554
|
|
Venn A A, Loram J E, Douglas A E. 2008. Photosynthetic symbioses in animals. Journal of Experimental Botany, 59(5): 1069–1080. doi: 10.1093/jxb/erm328
|
|
Ware J R, Smith S V, Reaka-Kudla M L. 1992. Coral reefs: sources or sinks of atmospheric CO2?. Coral Reefs, 11(3): 127–130
|
|
Weber L, Gonzalez-Díaz P, Armenteros M, et al. 2019. The coral ecosphere: a unique coral reef habitat that fosters coral–microbial interactions. Limnology and Oceanography, 64(6): 2373–2388. doi: 10.1002/lno.11190
|
|
Wiebe W J, Johannes R E, Webb K L. 1975. Nitrogen fixation in a coral reef community. Science, 188(4185): 257–259. doi: 10.1126/science.188.4185.257
|
|
Wild C, Huettel M, Klueter A, et al. 2004a. Coral mucus functions as an energy carrier and particle trap in the reef ecosystem. Nature, 428(6978): 66–70. doi: 10.1038/nature02344
|
|
Wild C, Niggl W, Naumann M S, et al. 2010. Organic matter release by Red Sea coral reef organisms—potential effects on microbial activity and in situ O2 availability. Marine Ecology Progress Series, 411: 61–71. doi: 10.3354/meps08653
|
|
Wild C, Rasheed M, Werner U, et al. 2004b. Degradation and mineralization of coral mucus in reef environments. Marine Ecology Progress Series, 267: 159–171. doi: 10.3354/meps267159
|
|
Wilkinson C R, Fay P. 1979. Nitrogen fixation in coral reef sponges with symbiotic cyanobacteria. Nature, 279(5713): 527–529. doi: 10.1038/279527a0
|
|
Williams I D, Baum J K, Heenan A, et al. 2015. Human, oceanographic and habitat drivers of central and western Pacific coral reef fish assemblages. PLoS ONE, 10(4): e0120516. doi: 10.1371/journal.pone.0120516
|
|
Williams J J, Papastamatiou Y P, Caselle J E, et al. 2018. Mobile marine predators: an understudied source of nutrients to coral reefs in an unfished atoll. Proceedings of the Royal Society B: Biological Sciences, 285(1875): 20172456. doi: 10.1098/rspb.2017.2456
|
|
Wu Chengye, Zhang Jianlin, Huang Liangmin. 2001. Primary productivity in some coral reef lagoons and their adjacent sea areas of Nansha Islands in spring. Journal of Tropical Oceanography (in Chinese), 20(3): 59–67
|
|
Wyatt A S J, Falter J L, Lowe R J, et al. 2012. Oceanographic forcing of nutrient uptake and release over a fringing coral reef. Limnology and Oceanography, 57(2): 401–419. doi: 10.4319/lo.2012.57.2.0401
|
|
Wyatt A S J, Leichter J J, Toth L T, et al. 2020. Heat accumulation on coral reefs mitigated by internal waves. Nature Geoscience, 13(1): 28–34. doi: 10.1038/s41561-019-0486-4
|
|
Wyatt A S J, Lowe R J, Humphries S, et al. 2010. Particulate nutrient fluxes over a fringing coral reef: relevant scales of phytoplankton production and mechanisms of supply. Marine Ecology Progress Series, 405: 113–130. doi: 10.3354/meps08508
|
|
Wyatt A S J, Lowe R J, Humphries S, et al. 2013. Particulate nutrient fluxes over a fringing coral reef: Source-sink dynamics inferred from carbon to nitrogen ratios and stable isotopes. Limnology and Oceanography, 58(1): 409–427. doi: 10.4319/lo.2013.58.1.0409
|
|
Xu Shendong, Zhang Zhinan, Yu Kefu, et al. 2021. Spatial variations in the trophic status of Favia palauensis corals in the South China Sea: insights into their different adaptabilities under contrasting environmental conditions. Science China: Earth Sciences, 64(6): 839–852. doi: 10.1007/s11430-020-9774-0
|
|
Yahel G, Post A F, Fabricius K, et al. 1998. Phytoplankton distribution and grazing near coral reefs. Limnology and Oceanography, 43(4): 551–563. doi: 10.4319/lo.1998.43.4.0551
|
|
Yamaguchi R, Suga T. 2019. Trend and variability in global upper-ocean stratification since the 1960s. Journal of Geophysical Research: Oceans, 124(12): 8933–8948. doi: 10.1029/2019JC015439
|
|
Yan Hongqiang, Yu Kefu, Shi Qi, et al. 2016. Seasonal variations of seawater pCO2 and sea-air CO2 fluxes in a fringing coral reef, northern South China Sea. Journal of Geophysical Research: Oceans, 121(1): 998–1008. doi: 10.1002/2015JC011484
|
|
Zaneveld J R, Burkepile D E, Shantz A A, et al. 2016. Overfishing and nutrient pollution interact with temperature to disrupt coral reefs down to microbial scales. Nature Communications, 7: 11833
|
|
Zhang Fan, Blasiak L C, Karolin J O, et al. 2015. Phosphorus sequestration in the form of polyphosphate by microbial symbionts in marine sponges. Proceedings of the National Academy of Sciences of the United States of America, 112(14): 4381–4386. doi: 10.1073/pnas.1423768112
|


Figures(2)

Supported by:
Beijing Renhe Information Technology Co. Ltd


| Water masses | ω | |||
| CEW | KW | YSCC | ||
| Salinity | 25.81 | 34.81 | 32.15 | 3.5 |
| Temperature/°C | 11.37 | 24.47 | 9.64 | 1.9 |
| 234U/238U activity ratio | 1.170 7 | 1.148 1 | 1.158 4 | 0.005 0 |
| Note: ω is the standard deviation of each property. CEW: estuarine water of Changjiang River, KW: Kuroshio water, YSCC: Yellow Sea Coastal Current. | ||||
DownLoad:
CSV
 DownLoad:
DownLoad:






 DownLoad:
DownLoad:
