

Figure
1.
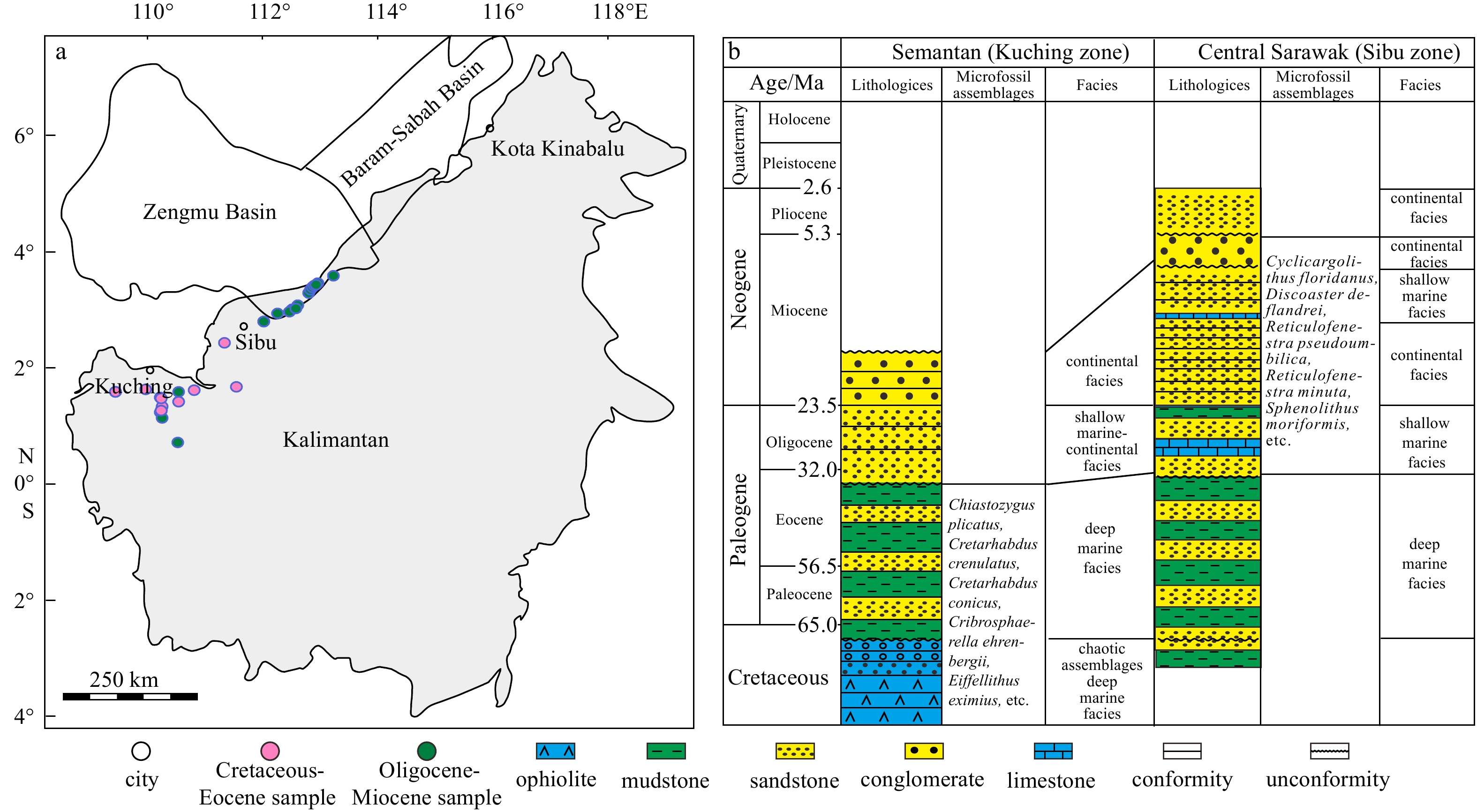
Locations of the outcrop samples in Kalimantan (a) and the generalized stratigraphic column of the Sarawak area (b) (modified after Wang et al. (2016)).
| Citation: | Lei Lan, Youchuan Li, Zhigang Zhao, Shuchun Yang, Qing Lin, Weilai Zhang. The influence of organic sources and environments on source rock deposition during the periods of Cretaceous–Eocene and Oligocene–Miocene, northern Kalimantan[J]. Acta Oceanologica Sinica, 2023, 42(3): 54-64. doi: 10.1007/s13131-022-2080-7

|
The South China Sea (SCS) covers an area of approximately 350×104 km2, and it has experienced two tectonic cycles, in which 14 large basins were formed and are mainly distributed on the continental shelf and slope (Zhang et al., 2018, 2021a). Until 2020, hundreds of gas fields had been found, 37 of which are large gas fields, and they are mainly distributed in the Yinggehai Basin, the Qiongdongnan Basin and the Pearl Mouth River Basin in the northern SCS, the Zengmu Basin and the Baram-Sabah Basin in the southern SCS with reserves more than 300×108 m3. These resources account for approximately 70% of the total gas reserves in the SCS.
As the most petroliferous basins in the southern SCS, the source rocks in the Zengmu Basin and the Baram–Sabah Basin have attracted enough attentions. According to earlier research, the source rocks in the Zengmu Basin and the Baram-Sabah Basin are the Oligocene and the Miocene strata which were deposited in the transitional facies (Lan, 2019; Li et al., 2021). These research have focused on the depositional environments and the hydrocarbon-generation potential of coal-measure source rocks (Wan Hasiah et al., 2013; Sia et al., 2014; Togunwa et al., 2015) and the controlling factors of organic matter accumulation (Li et al., 2021) but without thorough discussion on the biomarker signatures of the Oligocene and the Miocene shales and coal measures. Meanwhile, these Tertiary sediments were overlying the thick deep marine sediments of the Cretaceous–Eocene period. Previous studies on the Cretaceous–Eocene strata aimed to mark the sedimentary and stratigraphic architecture but the geochemical characteristics and hydrocarbon generation potential of the source rocks have received scant attention. Ahmed et al. (2021) has used Rock-Eval pyrolysis and total organic carbon analysis to evaluate the thermal maturity, organic matter richness and kerogen type of the Belaga Formation deposited during the late Cretaceous to the late Eocene. There is still no comprehensive studies have been carried out on the organic sources or hydrocarbon generation to discuss their possibility as source rocks.
The purpose of this study is to assess the geochemical variability of the Cretaceous–Eocene and the Oligocene–Miocene outcrop argillaceous rocks in the northwest of Kalimantan, to reveal the difference of the organic sources and hydrocarbon generation abilities of the source rocks in marine and transitional facies. Then, this paper tries to construct models for high quality marine source rock in the Cretaceous–Eocene period and high quality source rocks in transitional faicies in the Oligocene–Eocene period using palynological and geochemical data, and to provide references for petroleum resource evaluation and exploration in the southern SCS.
The SCS had experienced complicated tectonic activities (Zhang et al., 2021b). A unified basement of the SCS was initially formed in the early Cretaceous and then was separated into several sub-basement (Zhang et al., 2018). During the Cretaceous, with the closure of the Meso-Tethyan Ocean and the expansion of the neo-Tethyan Ocean, the western Kalimantan block and the eastern Kalimantan block drifted and collided (Hall, 2002). The Semitau Block in the Kuching zone in Kalimantan was separated from the southeast China continental margin, then drifted during the Paleocene, and was collided with the northern part of Kalimantan in the Eocene (Zahirovic et al., 2014). During the period of the late Eocene to the early Oligocene, the Zengmu Basin and the Baram-Sabah Basin collided with Borneo during the process of subduction and shrinking of the palaeo-SCS. This tectonic event resulted in the uplift and erosion of Borneo and the formation of the Zengmu Basin and the Baram-Sabah Basin as the compressional foreland basins (Wang et al., 2020). However, some research believed that the subduction of the palaeo-SCS started at Paleocene, or even at the late Cretaceous and the collisional event in the Zengmu Basin and Borneo started at the late Eocene (Zhang et al., 2018). During the upper Cretaceous to the middle Miocene, the terrestrial-marine siliciclastic sediments were deposited in the transitional part of the Zengmu Basin, where outcropping samples were obtained (Fig. 1a). After the middle Miocene, Kalimantan contra rotated and formed the current structural characteristics.
Controlled by the tectonic evolution from the Cretaceous to the middle Eocene, transgression occurred and there was a large scope of deep-sea area in the Sarawak and Sabah area of Kalimantan (Briais et al., 1993). The Kuching zone was overlain by deep-sea sediments of the Lupar Formation which was dated to the upper Cretaceous based on foraminiferal evidence (Tan, 1982). The Sibu area, which is located in the onshore part of the Zengmu Basin, was underlain predominantly by the Lupar and Belaga Formation. Foraminiferal dating placed the Belaga Formation in the upper Cretaceous to the upper Eocene. It forms a broad outcrop band that is almost 200 km wide and trends eastward in a crescentic pattern into the Sabah Basin (Berhad, 1999). Since the late Eocene, regression occurred and only the northeast area remained to be the deposition center of the marine system in the northwest of Kalimantan (Hall and Breitfeld, 2017). Since the early Oligocene, the uplift of Kalimantan has provided abundant material sources, which were beneficial for the development of two large deltas from the southern Zengmu. Sediments in the Baram-Sabah Basin was continuously deposited under deep-water environment. Until the early−middle Miocene, large-scale deltas were constantly developed in the Zengmu Basin and multiple delta deposits in a delta-neritic-abysmal sedimentary sequence began to fill the Baram-Sabah Basin (Guo et al., 2006).
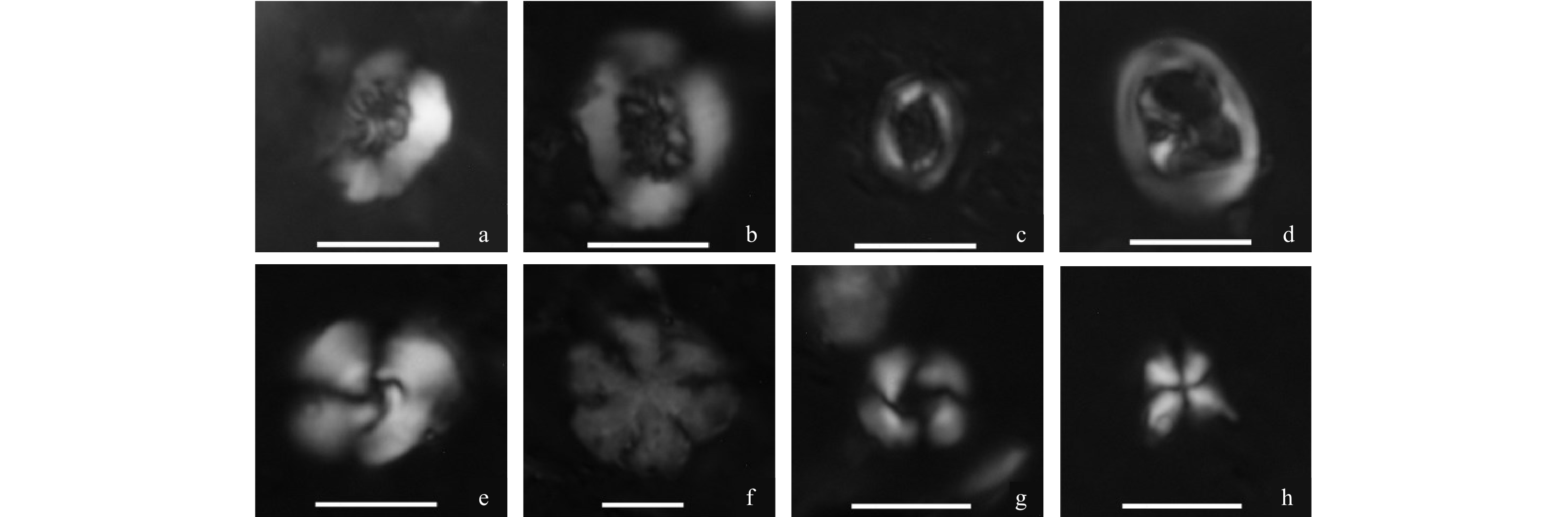
Outcrop samples obtained in Kalimantan is representative of various strata deposited from the Cretaceous to the Neogene in the Zengmu and the Baram-Sabah Basins. A total of thirty-six outcrop samples in different sedimentary facies were collected along the northwest margin of Kalimantan (Fig. 1). Age constraint of the outcrops is based on the geological map of Sarawak, calcareous nannofossils and sporopollen. Calcareous nannofossils were only found in K12, Q7, Q12, Q14 and Q16. Common nannofossils, widely distributed in the Cretaceous period, such as Chiastozygus plicatus, Cretarhabdus crenulatus, Cretarhabdus conicus, Cribrosphaerella ehrenbergii, Eiffellithus eximius, Eiffellithus turriseiffelii, etc. were detected in K12 samples (Fig. 2), while in other 4 samples, the appearance of the diagnostic nannofossils, such as Cyclicargolithus floridanus, Discoaster deflandrei, Reticulofenestra pseudoumbilica, Reticulofenestra minuta, Sphenolithus moriformis, etc. manifested the Oligocene–Miocene of stratigraphic age (Fig. 2). Although the majority of the observed spores and pollen were widely distributed in the Paleogene and the Neogene, there were still some pollen that had stratigraphic significance. Cicatricosisporites, Schizaeoisporites and Tricolpopollenites have been observed in Sample K12. Cicatricosisporites and Schizaeoisporites that were widely found in the early Cretaceous strata in Asia, suggested the deposition age of the Cretaceous–Eocene period. The appearance of Tricolpopollenites has also proved that the depositional time was later than the early Cretaceous period (OláníyìOdébòdé, 1987). Magnastriatites hawardtii was found in Samples Q5 and Q12. The first appearance of Magnastriatites hawardtii in Kalimantan was in the very late period of the Eocene and the continuous distributed from the Oligocene. Florschuetzia, an important marker genus in dating the Miocene sediments, especially within Kalimantan and adjacent areas (Morley, 1977; Lei, 1998), was found in Q7, Q9, Q15, Q16, Q20 and Q21 sample in high abundance. Therefore, samples from K1 to K12 and Q1 to Q4 were regarded as the sediments of the Cretaceous–Eocene period, K13 and K14 and Q5 to Q22 were deposited during the Oligocene to the Eocene period. Fteen of the collected samples are the Cretaceous to the Eocene shales in the Kuching zone from the Cretaceous to the Eocene strata and one is in the Sibu area. Lithologies of the other samples are coal, carbonaceous shales and shales that are mostly distributed in the north of Sibu in the Oligocene and the Miocene strata. Detailed information of these samples is provided in Table 1.
| Sample ID | Period/series | Facies | P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | P11 | P12 | P13 | P14 | P15 |
| K1 | C.–E. | B | –24.0 | 1.1 | 468 | 2.6 | 43 | 0.83 | 0.08 | 0.11 | 0.98 | 0.10 | 0.12 | 0.11 | 0.40 | 6.88 | 1.95 |
| K2 | C.–E. | B | –23.9 | 0.6 | 436 | 2.5 | 22 | N.A. | 0.07 | 0.72 | 1.04 | 0.11 | 0.12 | 0.13 | 0.58 | 8.79 | 2.06 |
| K3 | C.–E. | B | –23.8 | 0.5 | 449 | 1.7 | 31 | 0.83 | 0.09 | 0.75 | 0.85 | 0.14 | 0.24 | 0.10 | 0.29 | 13.00 | 1.95 |
| K4 | C.–E. | B | N.A. | 0.7 | 484 | 1.3 | 56 | N.A. | 0.07 | 0.14 | 0.39 | 0.27 | 0.46 | 0.16 | 0.88 | 6.09 | 1.96 |
| K5 | C.–E. | B | –22.0 | 0.1 | 593 | 2.1 | 3 | 1.47 | 0.06 | 0.06 | 0.25 | 0.41 | 0.80 | 0.14 | 0.23 | 6.87 | 1.74 |
| K6 | C.–E. | B | –23.5 | 0.1 | 588 | 2.7 | 5 | 1.12 | 0.08 | 0.00 | 0.39 | 0.40 | 0.71 | 0.15 | 0.31 | 5.99 | 1.86 |
| K7 | C.–E. | B | –24.6 | 0.1 | 369 | 0.9 | 8 | 2.27 | 0.07 | 0.04 | 0.34 | 0.47 | 0.74 | 0.15 | 0.25 | 5.27 | 1.64 |
| K8 | C.–E. | B | –24.8 | 0.0 | 477 | 0.8 | 4 | 3.61 | 0.05 | 0.21 | 0.36 | 0.43 | 0.82 | 0.15 | 0.34 | 5.05 | 1.74 |
| K9 | C.–E. | B | –26.2 | 0.1 | 597 | 1.5 | 5 | 2.42 | 0.07 | 0.04 | 0.25 | 0.51 | 0.77 | 0.15 | 0.19 | 4.74 | 1.52 |
| K10 | C.–E. | B | –26.7 | 0.1 | 596 | 2.6 | 4 | 2.26 | 0.07 | 0.03 | 0.36 | 0.43 | 0.73 | 0.14 | 0.22 | 4.71 | 1.77 |
| K11 | C.–E. | B | –27.3 | 0.1 | 598 | 3.0 | 4 | 2.57 | 0.07 | 0.05 | 0.38 | 0.33 | 0.57 | 0.17 | 0.38 | 5.15 | 1.40 |
| K12 | C.–E. | B | –24.6 | 0.2 | 448 | 0.6 | 30 | N.A. | 0.02 | 0.07 | 0.56 | 0.52 | 0.73 | 0.06 | 0.35 | 1.57 | 0.99 |
| Q1 | C.–E. | B | –27.2 | 0.0 | 576 | 0.8 | 4 | 1.50 | 0.07 | 0.01 | 0.36 | 0.60 | 1.11 | 0.12 | 0.34 | 4.52 | 1.54 |
| Q2 | C.–E. | B | –27.8 | 0.0 | 477 | 0.6 | 5 | 3.64 | 0.07 | 0.02 | 0.47 | 0.56 | 0.91 | 0.14 | 0.25 | 3.81 | 1.64 |
| Q3 | C.–E. | B | –27.9 | 0.3 | 461 | 0.4 | 77 | N.A. | 0.11 | 0.07 | 0.78 | 0.36 | 0.43 | 0.15 | 0.48 | 4.95 | 1.44 |
| Q4 | C.–E. | B | –26.8 | 0.1 | 597 | 0.5 | 10 | 2.66 | 0.06 | 0.02 | 0.54 | N.D. | N.D. | 0.12 | 0.44 | 4.65 | 1.54 |
| Q5 | O.–M. | B | –26.8 | 0.6 | 457 | 0.9 | 67 | N.A. | 0.46 | 8.72 | 2.53 | 1.48 | 0.57 | 0.11 | 0.48 | 4.40 | 0.28 |
| Q6 | O.–M. | B | –26.5 | 0.3 | 439 | 0.4 | 73 | N.A. | 0.16 | 2.12 | 2.43 | 2.53 | 0.98 | 0.10 | 0.68 | 0.71 | 0.76 |
| Q7 | O.–M. | B | –27.0 | 0.3 | 444 | 0.3 | 84 | 0.81 | 0.21 | 1.19 | 1.97 | 1.69 | 0.95 | 0.11 | 0.49 | 0.52 | 1.33 |
| Q8 | O.–M. | B | –28.4 | 0.3 | 415 | 1.0 | 29 | 0.79 | 0.66 | 1.21 | 1.27 | 1.29 | 1.12 | 0.05 | 0.35 | 2.68 | 0.77 |
| K13 | O.–M. | D-N | –26.0 | 1.0 | 527 | 3.2 | 33 | 0.95 | 0.13 | 0.17 | 0.12 | 0.37 | 0.76 | 0.11 | 0.26 | 2.40 | 1.51 |
| K14 | O.–M. | D-N | N.A. | 2.6 | 443 | 2.7 | 98 | N.A. | 0.21 | 3.35 | 3.10 | 2.11 | 0.62 | 0.05 | 0.39 | 1.44 | 0.47 |
| Q9 | O.–M. | D-N | –27.8 | 4.5 | 448 | 2.9 | 154 | 0.47 | 1.63 | 3.69 | 3.11 | 2.06 | 0.61 | 0.10 | 1.39 | 0.63 | 0.58 |
| Q10 | O.–M. | D-N | –27.0 | 0.4 | 433 | 0.4 | 120 | N.A. | 0.26 | 1.12 | 3.13 | 3.86 | 1.10 | 0.11 | 0.73 | 0.49 | 0.27 |
| Q11 | O.–M. | D-N | –26.5 | 0.2 | 435 | 0.3 | 70 | 1.66 | 0.08 | 1.36 | 1.39 | 1.34 | 0.89 | 0.07 | 0.68 | 0.94 | 0.78 |
| Q12 | O.–M. | D-N | –27.2 | 0.7 | 440 | 0.6 | 107 | 0.39 | 0.19 | 1.20 | 2.49 | 2.21 | 0.89 | 0.08 | 1.01 | 0.32 | 0.62 |
| Q13 | O.–M. | D-N | –27.5 | 0.8 | 442 | 0.7 | 109 | 0.49 | 0.19 | 1.99 | 2.65 | 2.12 | 0.87 | 0.07 | 0.75 | 0.36 | 0.62 |
| Q14 | O.–M. | D-N | –27.1 | 0.5 | 451 | 0.5 | 99 | 1.06 | 0.41 | 1.43 | 2.56 | 2.07 | 0.79 | 0.08 | 1.40 | 0.30 | 0.57 |
| Q15 | O.-M. | D-N | –27.1 | 1.2 | 411 | 1.7 | 75 | 0.65 | 4.39 | 1.21 | 11.98 | N.D. | 0.36 | 0.06 | 0.69 | 0.82 | 0.34 |
| Q16 | O.–M. | D-N | –28.3 | 1.0 | 441 | 0.7 | 133 | N.A. | 0.31 | 0.98 | 2.97 | 3.10 | 1.28 | 0.11 | 0.48 | 0.67 | 0.48 |
| Q17 | O.–M. | C | –27.4 | 29.8 | 445 | 18.7 | 159 | 0.61 | 0.14 | 0.11 | 4.55 | 1.09 | 0.20 | 0.07 | 3.47 | 0.39 | 0.77 |
| Q18 | O.–M. | C | –28.0 | 12.3 | 455 | 9.2 | 134 | 0.78 | 0.76 | 36.64 | 3.45 | 1.60 | 0.53 | 0.03 | 1.12 | 1.78 | 1.42 |
| Q19 | O.–M. | C | –27.2 | 145.7 | 441 | 53.3 | 273 | 0.71 | 0.13 | 0.80 | 3.66 | 3.14 | 1.35 | 0.05 | 3.10 | 0.09 | 0.89 |
| Q20 | O.–M. | C | –26.7 | 29.4 | 433 | 14.9 | 197 | 0.63 | 0.20 | 2.26 | 3.37 | 4.09 | 1.86 | 0.08 | 1.42 | 0.09 | 1.02 |
| Q21 | O.–M. | C | –27.9 | 31.9 | 426 | 21.1 | 152 | N.A. | 1.27 | 0.57 | 3.60 | 0.44 | 1.24 | 0.11 | 0.58 | 0.59 | 0.00 |
| Q22 | O.–M. | C | –26.8 | 1.8 | 412 | 7.3 | 25 | 0.40 | 2.03 | 0.80 | 2.54 | 2.82 | 1.68 | 0.04 | 0.16 | 3.59 | 0.27 |
| Note: C.–E.=Cretaceous–Eocene; O.–M.=Oligocene–Miocene; B=bathyal; N-D=delta-neritic; C=coaly; N.A.=not analyzed; N.D.=no data; P1=$ \text{δ}^{13} $Ckerogen; P2= S2, mg/g; P3=Tmax, ℃; P4=total organic carbon (TOC), %; P5=HI=hydrocarbon index, mg/g; P6=Ro, %; P7=Oleanane/C30 $\alpha \beta $ hopane; P8=bicadinane T/C30 $\alpha \beta $ hopane (m/z 412); P9=Pr/Ph; P10=Pr/nC17; P11=Ph/nC18; P12=Gam/C30 $\alpha \beta $ hopane; P13=(C19+C20)/(C23+C24) tricyclic terpanes; P14=(C19−C29) tricyclic terpanes/C30 $\alpha \beta $ hopane; P15=C27/C29 $\alpha \alpha \alpha $ 20R steranes. | |||||||||||||||||
 DownLoad:
CSV
DownLoad:
CSV
After removing external surfaces using a rock saw, the freshly-cut samples were washed with methanol and deionized water to remove the possible surficial contamination, the outcrop samples were then used for the following analyzing processes.
The samples were powdered to 200 mesh for Rock-Eval pyrolysis and total organic carbon (TOC) measurement. Rock-Eval pyrolysis was performed on a Rock-Eval VI to determine the amounts of free hydrocarbons (S1), potential hydrocarbons (S2), and the temperature of maximum generation (Tmax). The TOC was determined on a Leco CS-344 analyser, after pre-treatment of the samples with 5% dilute hydrochloric acid for 12 h and rinsing with deionised water 50 times to completely remove residual hydrochloric acid.
For the vitrinite reflectance (Ro) measurements, samples were crushed and embedded in liquid epoxy resin, polished to a smooth surface, and desiccated for 12 h. Ro was measured using a Leica MPV-SP microphotometer under an oil-immersion objective lens that was calibrated using a double calibration method using two Ro standards. The samples were re-measured if the measurement deviation of the standard samples was greater than ±0.02%. Between 35 to 50 vitrinite reflectance measurements were recorded for each sample, with the average Ro reported in Table 1.
The aliphatic hydrocarbons of the outcrop samples were obtained from solvent extraction and fractionation and then were analysed by gas chromatography-mass spectrometry (GC-MS) on an Agilent 6890 GC interfaced to an Agilent 5973 mass selective detector, with the ionisation source operated at an electron impact energy of 70 eV. An HP-5ms fused silica capillary column was used (30 m×0.25 mm i.d., film thicknesses 0.25 μm). The oven temperature was programmed to start at 20℃ (hold for 1 min), increased to 100℃ at 20℃/min, and from 100℃ to 310℃ at 3℃/min, with a final hold of 18 min. Helium was used as the carrier gas at a flow rate of 1.0 mL/min. Biomarker ratios were calculated based on their relative peak areas of individual compounds.
GC-MS-MS analyses were performed on a TSQ 8000 system (Thermo Fisher Scientific, USA). Samples were injected into the GC system using the AS 3000 auto-sampler. The GC was equipped with a HP-5ms used silica capillary column (60 m×0.25 mm i.d.×0.52 μm film thickness). Splitless mode was used with the inlet temperature set at 300℃, and the split flow at 15 mL/min following splitless time for 1.00 min. Helium (99.999%) was used as the carrier gas under constant flow mode at a rate of 1.0 mL/min. The GC oven temperature was initially set at 50℃ for 1 min, then programmed at 20℃/min to 120℃, then at 4℃/min to 250℃, then at 3℃/min to 300℃ and held for 30 min. The triple quadrupole mass spectrometer was operated in EI mode at 70 eV. The temperatures of the MS transfer line and ion source were set at 280℃ and 230℃, respectively. The filament emission current was 75 μA, and the filament-multiplier delay was set at 7 min. The tandem mass spectrometric detection was operated in the MRM mode for the highest possible selectivity and sensitivity. Collision-induced dissociation was performed using ultrapure nitrogen as the collision gas in the second quadrupole (Q2) collision cell. The first quadrupole (Q1) and third quadrupole (Q3) were tuned at unit mass resolution.
The sporopollen analyses were processed under the well-established Chinese industrial standard procedure for pollen extraction (SY/T 5522-2018), using 10% hydrochloric acid to eliminate carbonates and 40% hydrofluoric acid to eliminate silicates. A heavy liquid (HBr, KI, Zn) with a specific gravity of 2.0 (d=2) was used to separate the pollen from the remaining inorganic fraction. The palynological residues were sieved using 10 μm mesh nylon sieves in an ultrasonic bath. Sporopollen were identified and counted on a binocular microscope (Leica DM4000B) using 400×magnification. An average of 38 and 84 pollen grains were counted respectively in K samples and Q samples. The proportion of individual sporopollen taxa were determined relative to the total amount of spore and pollen (%).
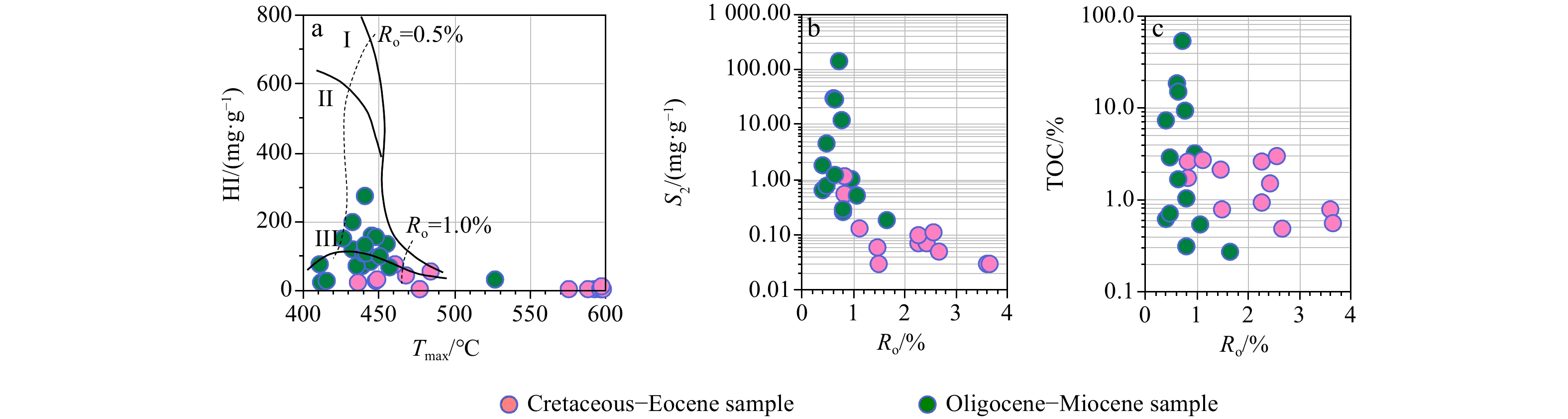
The Cretaceous–Eocene Formation samples have total organic carbon (TOC) contents of 0.5%–3.0% (average 1.54%), Rock-Eval S2 values (the content of HC in rock) of 0.03–1.10 mg /g (average 0.3 mg /g), hydrogen indices of 6–77 mg /g (average 20.5 mg /g) and vitrinite reflectance of 0.65%–3.64% (Table 1, Fig. 3). The positive correlation between Ro and S2 in the Cretaceous–Eocene samples suggests that the high thermal evolution degree is associated with the decrease in S2. The cross-plot of Ro and TOC indicates that the high maturity had minor influence on TOC value, proving that TOC can be used to reflect the hydrocarbon generation potential of the source rocks. The TOC contents of the Oligocene–Miocene Formation samples range from 0.3% to 53.3% (average (the content of HC in rock) 7.0%), Rock-Eval S2 values range from 0.2 mg /g to 145.7 mg /g (average 13.3 mg /g), hydrogen indices range from 25 mg /g to 273 mg /g (average 109.5 mg /g), and vitrinite reflectance ranges from 0.39% to 1.65%. Most of the Oligocene–Miocene samples have Ro lower than 1.2%, suggesting that the samples are in the immature to peak stages of hydrocarbon generation. The relative low values of hydrogen indices suggest that the organic matters are dominated by terrestrial higher plants.
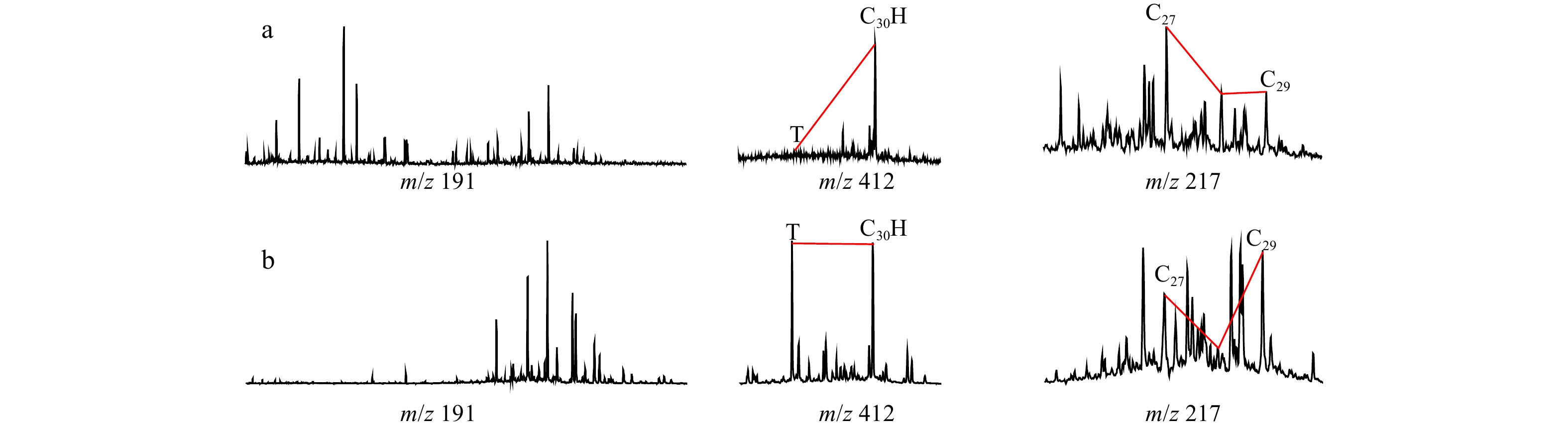
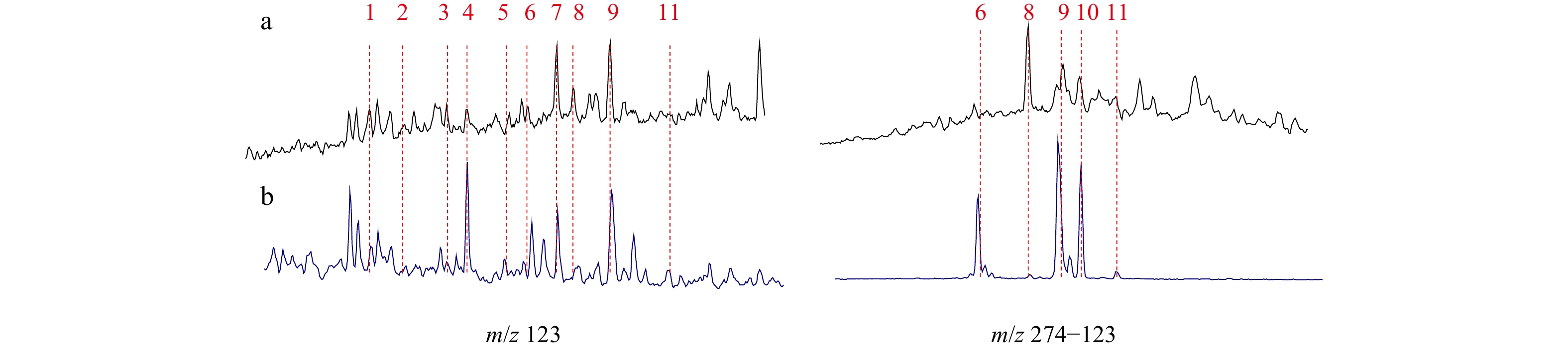
Aliphatic hydrocarbons of the outcropping samples were analyzed by GC-MS. Representative m/z 191, m/z 412 and m/z 217 mass chromatograms for the analyzed outcrops are shown in Fig. 4. All the samples have various distributions of tricyclic terpanes, hopanes and steranes (Table 1), suggesting obvious different origins and/or depositional environments for the organic matter in the samples. Twenty-eight samples were selected for GC-MS-MS analysis to further identify the diterpanes in aliphatic fraction (Fig. 5). Due to their low abundances, peak areas and the related diterpane parameters are not calculated. Nevertheless, the distinct patterns between the Cretaceous–Eocene and the Oligocene–Miocene samples provide evidence of the original discrepancy of the organic matter in source rocks.
Gymnosperm pollen are the dominant in flora, with an average proportion of 63% in the Cretaceous–Eocene samples. Fern spores have the average proportion of 32% in the coeval palynological records. In contrast, there are less angiosperms and algae, with the proportion of 3% and 2% respectively. Gymnosperm pollen are mainly dominated by Cheirolepidiaceae (42%) and Cupressaceae (21%). Cheirolepidiaceae are mainly composed of Classopollis annulatus. Exesipollenites tumulus and E. pseudotriletes are the two primary species of the Cupressaceae family. In addition, Abietineaepollenites, Pinuspollenites, Taxodiaceaepollenites hiatus, Ephedripites, and Cycadopites nitidus appeared sporadically. Fern spore types are characterized by Deltoidospora and Cyathidites minor, accounting for nearly 19% of all the spores and pollen. 8% of the sporepollen are Lygodiaceae spores, mainly Cicatricosisporites, Toroisporis and Klukisporites. Schizaeoisporites, Lygodiumsporites pseudomaximus, and other species make up 5% of the total (Fig. 6a).
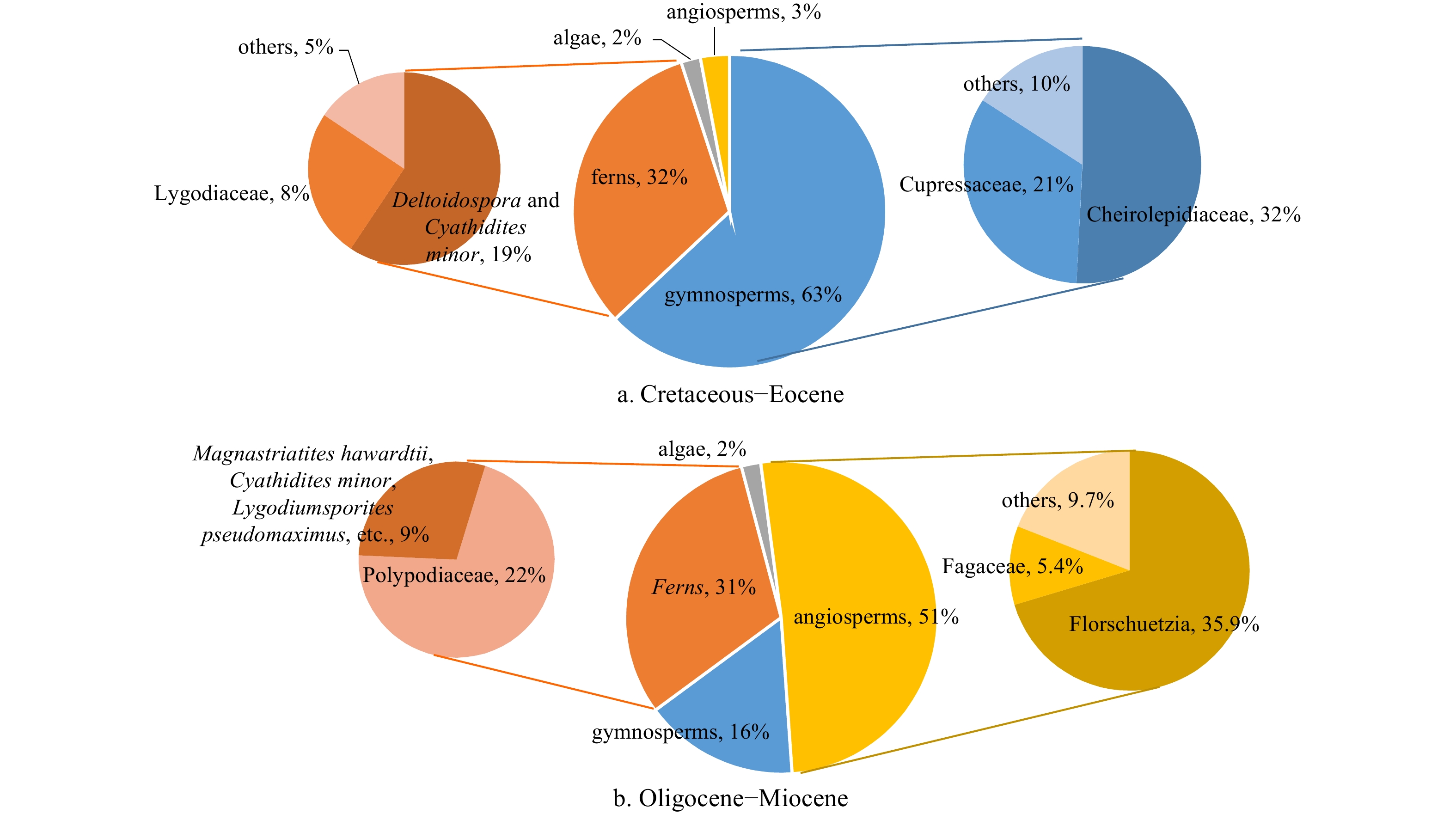
The sporopollen in the palynological records from the Oligocene and the Miocene outcrops include 51% of angiosperms, 31% of ferns, and 16% of gymnosperms (Fig. 6b). In the pollen of angiosperms, Florschuetzia, the Moraceae pollen, accounts for 35.9% of the total sporopollen fossil, followed by Fagaceae at 5.4%. Other angiosperms speices are mainly Tiliaepollenites, Betulaceoipollenites, Alnipollenites verus, Caryapollenites triangulus, Liquidambarpollenites, etc., accounting for 9.7%. The pollen species of gymnosperms are monotonous. Most of them are Pinaceae (Abietineaepollenites, Pinuspollenites, Tsugaepollenites igniculus, etc.), with a proportion of 14%. Polypodiaceae is the dominant species in Ferns, with the abundance of 22%. Other ferns speices are mainly Magnastriatites hawardtii, Cyathidites minor, and Lygodiumsporites pseudomaximus.
Cheirolepidiaceae, an extinct taxa of conifers, was found in records of spores and pollen during the Cretaceous and the Eocene, and its thriving is suggestive of the arid and hot climate (Axsmith and Jacobs, 2005; Watson, 1988). Schizaeoisporites, one taxa of herbaceous fern, have been mainly distributed in a tropic and dry area in the southern hemisphere (Umetsu and Matsuoka, 2003; Umetsu and Sato, 2007). Ephedripites, a kind of dwarf shrub, are common in the dessert, arid steppe and saline lakeside zone (Hoorn et al., 2012). Cicatricosisporites and Klukisporites are herbaceous ferns and have a preferential adaptation to the tropical and subtropical environments (Ding and Zhang, 2004; Li and Liu, 1994; Zhong et al., 2012). These floral characteristics, taken as a whole, suggest an arid and tropical or subtropical environment throughout the Cretaceous–Eocene epoch (Fig. 7).

The Oligocene–Miocene palynological records during the Oligocene and the Miocene are quite different from the Cretaceous–Eocene spore and pollen. The Florschuetzia group pollen, which were identified in the samples, including F.semilobata, F. trilobat and F.levipoli, are derived from the mangrove genus Sonneratia which is philotherm (Mao and Foong, 2013; Mustapha et al., 2017; Zhou and Lin, 2001). Magnastriatites hawardi are diagnostic spores of ferns, have the ecologically preferential adaptation to the tropical and subtropical climate (Mkpong et al., 2019). According to the sporepollen compositions, the Oligocene and the Miocene epochs were marked by a steady warming and humidification of the climate, which culminated in a tropical setting. This conclusion is in line with previous work from Morley (2018).
Gammacerane is believed to be formed by the reduction of tetrahymanol, hence the gammacerane/C30 αβ hopane (Gam/C30H) is used to indicate the stability of the stratified water column and perhaps water salinity in marine and non-marine source rock depositional environments (Damsté et al., 1995). Relative abundance of pristine/phytane (Pr/Ph), Pr/nC17 and Ph/nC18 have been widely used to evaluate the depositional environments of the organic matter (Didyk et al., 1978; Shanmugam, 1985). There was an obvious variation of sedimentary conditions from the Cretaceous–Eocene to the Oligocene–Miocene periods. In comparison to samples in other ages (Fig. 8a), the Cretaceous-Eocene samples show the lowest Pr/Ph ratios and the greatest Gam/C30H ratios (>0.1), which suggests that anoxic and saline water conditions were more prevalent in the more still water during this time. It is consistent with previous study in which the restricted water was caused by the weakening influence of the palaeo-Pacific on the palaeo-SCS and weak interaction between the palaeo-Pacific and the Tethys Ocean in the Cretaceous (Zhang et al., 2018). In contrast, the Oligocene–Miocene samples have the highest Pr/Ph ratios and the lowest Gam/C30H ratios, reflecting less reducing and larger influx of freshwater during deposition of the Oligocene–Miocene sediments. Hence, the water body in the shallow sea is constantly disturbed due to the adequate influx of terrestrial materials from delta to sea, causing the more oxidizing condition of the water environment. Additionally, the cross-plot of Ph/nC18 versus Pr/nC17 demonstrates that the majority of the Cretaceous–Eocene samples fall in the reducing district with a greater contribution from marine algae, whilst the Oligocene and the Miocene samples are deposited in a reducing to oxidizing environment, with most of them falling in the oxic zone (Fig. 8b).

Sterane parameters are effective to reflect primary producers in marine and lacustrine systems (Knoll et al., 2007). C27 steranes are believed to be originated from phytoplankton and metazoan, C29 steranes are mainly derived from terrigenous higher plants (Volkman, 1986). C27/C29 sterane ratios have high values (>1.5) in the Cretaceous–Eocene samples and exhibit low values (<1.5) in the Oligocene–Miocene samples (Table 1, Fig. 4). This suggests that the Cretaceous–Eocene to Oligocene–Miocene organic matter received reduced contribution from algae. Additionally, there is a positive correlation between (C19–C29) tricylic terpanes/C30 αβ hopane and C27/C29 ααα 20R steranes, which is indicative of a greater contribution from algae in the period of Cretaceous to Eocene compared to the period of the Oligocene and the Miocene (Fig. 8c).
Tricyclic terpanes are widespread in oils and source rock extracts. The extremely high tricyclic terpanes/C17α-hopanes is a source parameter that assesses the relative contribution from a group of bacterial or algal lipids (tricyclics) and various prokaryotic species (hopanes) (Peters et al., 2005). The patterns of tricyclic terpanes have a distinguishing difference between the Cretaceous–Eocene and Oligocene–Miocene samples. The Cretaceous–Eocene samples have a pronounced high content of tricyclic terpanes, with almost similar peak heights with pentacyclic triterpanes (Fig. 4a) compared to low relative abundance of tricylic terpanes in the Oligocene–Miocene samples (Fig. 4b). Some C19 and C20 tricyclic terpanes were thought to be derived from higher plants including gymnosperms and C23 tricyclic terpanes were in association with a possible marine algae source (Peters et al., 2005). The Cretaceous–Eocene samples have low (C19+C20) TT/(C23+C24) TT and high (C19–C29) TT/C30H compared to the Oligocene–Miocene samples (Fig. 8d), respectively, which supports the theory that higher plant-derived C19 and C20 tricyclic terpanes are not the causes of the high (C19+C20) TT/(C23+C24) TT ratio. Hence, such high ratios can reflect the major contribution of algae (probably Tasmanites algae) during the Cretaceous–Eocene period. In contrast, the Oligocene-Miocene samples have relatively lower ratios of (C19–C29) TT/C30H (mostly<1.6) but higher (C19+C20) TT/(C23+C24) TT than the younger samples, indicating large algal contributions of C19 and C20 tricyclic terpanes to the C19–C29 tricyclic terpanes. It further suggests the limited contribution from algae to the Oligocene–Miocene samples.
Abundance of tricyclic terpanes in Tasmanites rock extracts indicates a possible tricylic terpane origin from Tasmanites algae (Azevedo et al., 1992). Tasmanites alage is an important organic contributor in good-excellent quality source rocks, these algae are unexpectedly enriched in 13C (Peters et al., 2005). In addition, Tasmanites algae are known to have adaptation in a variety of environments with various water salinity and temperature in large range of geological periods (Guy-Ohlson, 1988). Blooms of Tasmanites alage happen two or three times annually in the western British Channel (Vigran et al., 2008). In the cross-plot of δ13Ckerogen versus (C19–C29) TT/C30 αβ hopane, samples with (C19–C29) TT/C30H>6 have heavier kerogen carbon isotopes (δ13Ckerogen>–24‰), while samples with (C19–C29) TT/C30 αβ hopane between 4 and 6 have lighter carbon isotopes of kerogen (most δ13Ckerogen<–26‰) (Fig. 8e). The samples with high (C19–C29) TT/C30H and heavy 13Ckerogen may shed light on the phenomenon that Tasmanites or other types of prasinophyte green algae that are similar to Tasmanites intermittently dominated the algal population as the primary producers during the Cretaceous–Eocene period. When the particular planktonic algae in the marine ecosystem declined, other planktonic algae with less 13C, such as blue-green algae and diatoms, were probably the main producers.
Angiosperms did not appear until the early-Cretaceous (Burger, 1980). Gymnosperms and ferns were still the dominant vegetation types during the Cretaceous (Dettmann et al., 1992). In study area, the gymnosperm-derived diterpanes were presented in very low abundance in all samples. Nevertheless, distribution of the detectable deterpanes include β-labdane, 19-norisopimarane, 18-norisopimarane, rimuane, 17-nortetracyclane, beyerane, isopimarane, β-phyllocladane, β-kaurane and ent-kaurane were classified into two patterns (Fig. 5). The Cretaceous–Eocene samples have similar patterns with high abundance of isopimarane, β-phyllocladane and β-kaurane, while the Oligocene–Miocene samples are characterized by relatively higher contents of rimuane, beyerane, isopimarane, β-kaurane and Atisane. Rimuane was originated from rimuene which was formed from the epicuticular wax and was rich in Cupressaceae, Taxodiaceae, Podocarpaceae as well as Phyllocladaceae (Salasoo, 1984). Beyeranes has been found in several species of conifers, including Cupressaceae, Araucariaceae and Podocarpaceae (Jiang and George, 2018). Isopimaranes were reported in all conifer communities (Otto and Wilde, 2001). β-phyllocladane was identified in resins from Podocarpus and Dacrydium (family of Podocarpaceae). Kaurane was detected in Agathis, a family of Araucariaceae (Killops et al., 1995). Unlike other kinds of diterpanes, atisane was isolated from the bark of the mangrove plant Excoecaria agallocha which was angiosperm-derived (Wang et al., 2009). Thus, these diterpanes, such as rimuane, beyerane, isopimarane and kaurane, which were chiefly rich in conifer resins, indicate the contribution of gymnosperms to the organic matter during the Cretaceous–Eocene and the Oligocene–Miocene periods. The difference in the relative abundance of rimuane and phyllocladane is likely due to the thermal evolution of the organic matter (Jiang and George, 2018). The low abundance of gymnosperm-derived diterpanes in the Cretaceous–Eocene samples was in accordance with the bathyal-abysmal sea environment, which was interpreted to have received limited supplies of terrigenous organic matter. In addition, the low abundance of gymnosperm-derived diterpanes in the Oligocene–Miocene samples is suggestive of the subordinate contribution of gymnosperm to the organic matter. The relatively abundant atisane detected in the Oligocene and the Miocene samples illustrates the increase in contribution of angiosperms, especially mangroves, compared to the Cretaceous–Eocene samples.
Globally, angiosperms became the dominant type in flora from the late Cretaceous. Oleanane, originated from betulins, taraxerene and other pentacyclic triterpenoids in angiosperms, is highly specific for angiosperm plant inputs during the Cretaceous or younger age (Peters et al., 2005). Bicadinanes are mostly derived from a resinous polymer produced by angiosperms such as the tropical/subtropical plants Dipterocarpaceae (van Aarssen et al., 1992). In recent years, many authors have reported abundant oleanane and bicadinanes in crude oils, source rocks and outcrops in the SCS and South East Asia (Wan Hasiah, 1999; Sia et al., 2014). High abundance of bicadinanes is considered to be associated with mangroves (Lan, 2019). As shown in Fig. 8f, lowest oleanane/C30 hopane and lowest T-bicadinane/C30hopane (T/C30H) ratios in the Cretaceous–Eocene samples indicate minor angiosperm contribution during this period. In contrast, the Oligocene–Miocene samples were plotted in the area with high Ol/C30H and T/C30H ratios. High abundance of oleanane, bicadinane and the negative correlation between Ol/C30H and C27/C29 ααα 20R (Fig. 8g) suggest that angiosperms were dominant types in flora. Abundant bicadinanes in the samples are also consistent with the presence of atisane in the Oligocene–Miocene samples.
In conclusion, despite gymnosperms being the main flora type during the Cretaceous–Eocene period, their contribution to organic matter was relatively low due to the lengthy onshore distance where scant terrigenous organic matter was deposited. Angiosperms contributed more organic matter than gymnosperms throughout the Oligocene–Miocene epoch.
Two models for source rock developments were constructed from the Cretaceous–Eocene period to the Oligocene–Miocene period.
During the Cretaceous–Eocene period, gymnosperms flourished. Due to the distance offshore, the organic matter that is received in the bathyal-abysmal marine is limited, and the majority of the organic matter is sea-derived algae. Tasmanites or other varieties of prasinophyte green algae may occasionally bloom and offer a rather high productivity. This can be substantiated by the strong positive correlations between (C19–C29) TT/C30H ratios and TOC, and between kerogen carbon isotopes and TOC (Figs 8h–i). Additionally, the good reduction of seawater allows the preservation of algae, which can be dedaced from the high C27/C29 content and low Pr/Ph of the Cretaceous–Eocene sample in Fig. 8j, which in turn allows the development of high-quality source rocks of the oil-prone.
Angiosperms dominated higher plants during the Oligocene–Miocene epoch, and their growth was facilitated by the humid climate. Mangroves that were hydrogen-rich served as a significant source of higher plant parent material in the deltaic-shallow marine sedimentary system. The coal developed in the delta with such organic parent material has a significantly stronger potential to produce oil than normal coal. In the shallow sea, the stronger hydrodynamic force will accelerate the dilution effect on land-derived organic matter and the enhanced oxidation of water body, which promote up the oxidation and degradation of the hydrogen-rich components in sea-derived algae and land-derived organic matter. As a result, the shallow marine source rocks from the Oligocene and the Miocene are gas-prone.
(1) The distinguishing biomarker assemblages of the 36 outcrop samples suggest the differences of organic matter inputs and preservation conditions of the Cretaceous–Eocene and the Oligocene–Miocene source rocks. Due to the weakened impact of the palaeo-Pacific on the palaeo SCS and the weakening connection between the palaeo-Pacific and Tethys Ocean, the sea was semi-closed and the water was salty and reducing over the Cretaceous-Eocene period. In addition, phytoplankton algae were the important contributors to the organic matter communities. During the Oligocene–Miocene, large-scale deltas developed forwards into the sea caused by the Sarawak orogeny. Strong hydrodynamic force in the shallow sea was not beneficial for the preservation of organic matter. Abundant angiosperms and terrestrial organic matter were carried into the delta, shallow sea or even deep sea environments. Higher plants became the dominant organic matter source compared to algae.
(2) The moderate to high primary productivity and reducing water conditions were beneficial for the formation of high-quality marine source rocks during the Cretaceous–Eocene period, although the low Rock-Eval S2 values and low hydrogen indices of the source rocks which were influenced by high thermal maturity. Abundant angiosperm-derived terrestrial organic matter, especially hydrogen-rich mangrove-origin plant materials, contributed to the formation of high-quality source rocks in marine-continental transitional facies during the Oligocene–Miocene period.
Acknowledgements: We appreciate the collaboration and enthusiastic support of Lei Shao from Tongji University for leading our expedition to Kalimantan for outcrops. We also appreciate the paleontological assistance from Youhua Zhu from Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences; Gongcheng Zhang, Wu Tang, and Yibo Wang from China National Offshore Oil Corporation (CNOOC) Research Institute. Dujie Hou from China University of Geosciences (Beijing), and Long Su from Oil and Gas Research Center, Northwest Institute of Eco-Environment and Resources, CAS are also thanked for their critical reviews and suggestions to improve the manuscript and figures. Wenjing Ding from Macquarie University in Australia was thanked for the language polishing.|
Ahmed N, Siddiqui N A, Rahman A H B A, et al. 2021. Evaluation of hydrocarbon source rock potential: deep marine shales of Belaga Formation of late Cretaceous-late Eocene, Sarawak, Malaysia. Journal of King Saud University-Science, 33(1): 101268. doi: 10.1016/j.jksus.2020.101268
|
|
Axsmith B J, Jacobs B F. 2005. The conifer Frenelopsis ramosissima (cheirolepidiaceae) in the lower Cretaceous of Texas: systematic, biogeographical, and paleoecological implications. International Journal of Plant Sciences, 166(2): 327–337. doi: 10.1086/427202
|
|
Azevedo D A, Aquino Neto F R, Simoneit B R T, et al. 1992. Novel series of tricyclic aromatic terpanes characterized in Tasmanian tasmanite. Organic Geochemistry, 18(1): 9–16. doi: 10.1016/0146-6380(92)90138-N
|
|
Berhad P N. 1999. The Petroleum Geology and Resources of Malaysia. Kuala Lumpur, Malaysia: Petroliam Nasional Berhad
|
|
Briais A, Patriat P, Tapponnier P. 1993. Updated interpretation of magnetic anomalies and seafloor spreading stages in the South China Sea: implications for the Tertiary tectonics of Southeast Asia. Journal of Geophysical Research: Solid Earth, 98(B4): 6299–6328. doi: 10.1029/92JB02280
|
|
Burger D. 1980. Palynological studies in the Lower Cretaceous of the Surat Basin, Australia. Canberra: Australian Government Publishing, 70–75
|
|
Damsté J S S, Kenig F, Koopmans M P, et al. 1995. Evidence for gammacerane as an indicator of water column stratification. Geochimica et Cosmochimica Acta, 59(9): 1895–1900. doi: 10.1016/0016-7037(95)00073-9
|
|
Dettmann M E, Molnar R E, Douglas J G, et al. 1992. Australian Cretaceous terrestrial faunas and floras: biostratigraphic and biogeographic implications. Cretaceous Research, 13(3): 207–262. doi: 10.1016/0195-6671(92)90001-7
|
|
Didyk B M, Simoneit B R T, Brassell S C, et al. 1978. Organic geochemical indicators of palaeoenvironmental conditions of sedimentation. Nature, 272(5650): 216–222. doi: 10.1038/272216a0
|
|
Ding Qiuhong, Zhang Lidong. 2004. Spore-pollen flora as the indicator of paleoclimate condition in the Yixian Formation, western Liaoning Province. Acta Micropalaeontologica Sinica, 21(3): 332–341
|
|
Guo Xiurong, Wu Qiang, Qiu Yan, et al. 2006. Analysis of the shelf-margin delta in the south of Zengmu Basin, South China Sea. Marine Geology & Quaternary Geology, 26(4): 1–6
|
|
Guy-Ohlson D. 1988. Developmental stages in the life cycle of Mesozoic Tasmanites. Botanica Marina, 31(5): 447–456
|
|
Hall R. 2002. Cenozoic geological and plate tectonic evolution of SE Asia and the SW Pacific: computer-based reconstructions, model and animations. Journal of Asian Earth Sciences, 20(4): 353–431. doi: 10.1016/S1367-9120(01)00069-4
|
|
Hall R, Breitfeld H T. 2017. Nature and demise of the Proto-South China Sea. Bulletin of the Geological Society of Malaysia, 63: 61–76. doi: 10.7186/bgsm63201703
|
|
Hoorn C, Straathof J, Abels H A, et al. 2012. A late Eocene palynological record of climate change and Tibetan Plateau uplift (Xining Basin, China). Palaeogeography, Palaeoclimatology, Palaeoecology, 344–345: 16–38
|
|
Jiang Lian, George S C. 2018. Biomarker signatures of upper Cretaceous Latrobe Group hydrocarbon source rocks, Gippsland Basin, Australia: distribution and palaeoenvironment significance of aliphatic hydrocarbons. International Journal of Coal Geology, 196: 29–42. doi: 10.1016/j.coal.2018.06.025
|
|
Killops S D, Raine J I, Woolhouse A D, et al. 1995. Chemostratigraphic evidence of higher-plant evolution in the Taranaki Basin, New Zealand. Organic Geochemistry, 23(5): 429–445. doi: 10.1016/0146-6380(95)00019-B
|
|
Knoll A H, Summons R E, Waldbauer J R, et al. 2007. The geological succession of primary producers in the oceans. In: Falkowski P G, Knoll A H, eds. Evolution of Primary Producers in the Sea. Boston: Academic Press, 133–163
|
|
Lan Lei. 2019. Controlling factors for different hydrocarbon distribution in basins in southern South China Sea. Geological Science and Technology Information, 38(4): 23–29
|
|
Lei Zuoqi. 1998. The tertiary and the distribution regularity of mangrove sporopollen in the Pearl River Mouth Basin. Guangdong Geology, 13(2): 49–54
|
|
Li Wenben, Liu Zhaosheng. 1994. The cretaceous palynofloras and their bearing on stratigraphic correlation in China. Cretaceous Research, 15(3): 333–365. doi: 10.1006/cres.1994.1021
|
|
Li Youchuan, Zhao Zhigang, Lan Lei, et al. 2021. Differential distribution of oil and gas in the Zengmu Basin and the Brunei-Sabah Basin under the joint control of source rock and thermal evolution degree. Natural Gas Industry, 41(11): 24–32
|
|
Mao Limi, Foong S Y. 2013. Tracing ancestral biogeography of Sonneratia based on fossil pollen and their probable modern analogues. Palaeoworld, 22(3/4): 133–143
|
|
Mkpong E O, Nnakenyi N I, Essien A E, et al. 2019. The occurrences of belskipollis elegans and magnastriatites howardi: a review of their usage for zonation in the middle Miocene of the Niger Delta. Journal of Scientific and Engineering Research, 6(10): 199–208
|
|
Morley R J. 1977. Palynology of tertiary and quaternary sediments in Southeast Asia. In: 6th Annual Convention Proceedings. Jakarta:Indonesian Petroleum Association, 255–276
|
|
Morley R J. 2018. Assembly and division of the South and South-East Asian flora in relation to tectonics and climate change. Journal of Tropical Ecology, 34(4): 209–234. doi: 10.1017/S0266467418000202
|
|
Mustapha K A, Abdullah W H, Konjing Z, et al. 2017. Organic geochemistry and palynology of coals and coal-bearing mangrove sediments of the Neogene Sandakan Formation, Northeast Sabah, Malaysia. CATENA, 158: 30–45. doi: 10.1016/j.catena.2017.06.005
|
|
OláníyìOdébòdé M. 1987. Palynological dating of the Lamja Sandstone (Benue Basin, Nigeria) and its geological significance. Journal of African Earth Sciences (1983), 6(4): 421–426. doi: 10.1016/0899-5362(87)90085-6
|
|
Otto A, Wilde V. 2001. Sesqui-, Di-, and triterpenoids as chemosystematic markers in extant conifers—a review. The Botanical Review, 67(2): 141–238. doi: 10.1007/BF02858076
|
|
Peters K E, Walters C C, Moldowan J M. 2005. The Biomarker Guide: Biomarkers and Isotopes in Petroleum Exploration and Earth History. 2nd ed. New York: Cambridge University Press
|
|
Salasoo I. 1984. Structure analysis of rimuene by 13C nmr spectroscopy. Phytochemistry, 23(1): 192–193. doi: 10.1016/0031-9422(84)83110-6
|
|
Shanmugam G. 1985. Significance of coniferous rain forests and related organic matter in generating commercial quantities of oil, Gippsland Basin, Australia. AAPG Bulletin, 69(8): 1241–1254
|
|
Sia S G, Abdullah W H, Konjing Z, et al. 2014. The age, palaeoclimate, palaeovegetation, coal seam architecture/mire types, paleodepositional environments and thermal maturity of syn-collision paralic coal from Mukah, Sarawak, Malaysia. Journal of Asian Earth Sciences, 81: 1–19. doi: 10.1016/j.jseaes.2013.11.014
|
|
Tan D N K. 1982. The Lubok Antu Melange, Lupar valley, West Sarawak: a lower tertiary subduction complex. Bulletin of the Geological Society of Malaysia, 15: 31–46. doi: 10.7186/bgsm15198204
|
|
Togunwa O S, Abdullah W H, Hakimi M H, et al. 2015. Organic geochemical and petrographic characteristics of Neogene organic-rich sediments from the onshore West Baram Delta Province, Sarawak Basin: implications for source rocks and hydrocarbon generation potential. Marine and Petroleum Geology, 63: 115–126. doi: 10.1016/j.marpetgeo.2015.02.032
|
|
Umetsu K, Matsuoka A. 2003. Early Cretaceous fossil spores and pollen from the Tetori Group in the upper reaches of the Kuzuryu River, Fukui Prefecture, central Japan. The Journal of the Geological Society of Japan, 109(7): 420–423. doi: 10.5575/geosoc.109.420
|
|
Umetsu K, Sato Y. 2007. Early Cretaceous terrestrial palynomorph assemblages from the Miyako and Tetori Groups, Japan, and their implication to paleophytogeographic provinces. Review of Palaeobotany and Palynology, 144(1–2): 13–24
|
|
van Aarssen B G K, Hessels J K C, Abbink O A, et al. 1992. The occurrence of polycyclic sesqui-, tri-, and oligoterpenoids derived from a resinous polymeric cadinene in crude oils from Southeast Asia. Geochimica et Cosmochimica Acta, 56(3): 1231–1246. doi: 10.1016/0016-7037(92)90059-R
|
|
Vigran J O, Mørk A, Forsberg A W, et al. 2008. Tasmanites algae—contributors to the middle Triassic hydrocarbon source rocks of Svalbard and the Barents Shelf. Polar Research, 27(3): 360–371. doi: 10.1111/j.1751-8369.2008.00084.x
|
|
Volkman J K. 1986. A review of sterol markers for marine and terrigenous organic matter. Organic Geochemistry, 9(2): 83–99. doi: 10.1016/0146-6380(86)90089-6
|
|
Wan Hasiah A. 1999. Oil-generating potential of Tertiary coals and other organic-rich sediments of the Nyalau Formation, onshore Sarawak. Journal of Asian Earth Sciences, 17(1–2): 255–267
|
|
Wan Hasiah A, Lee C P, Gou P, et al. 2013. Coal-bearing strata of Labuan: mode of occurrences, organic petrographic characteristics and stratigraphic associations. Journal of Asian Earth Sciences, 76: 334–345. doi: 10.1016/j.jseaes.2013.05.017
|
|
Wang Pengcheng, Li Su, Guo Lingli, et al. 2016. Mesozoic and Cenozoic accretionary orogenic processes in Borneo and their mechanisms. Geological Journal, 51(S1): 464–489
|
|
Wang Yibo, Zhao Zhigang, Xie Xiaojun, et al. 2020. Cenozoic sedimentary fillings and development characteristics of source rocks in main basins in central-southern South China Sea. China Offshore Oil and Gas, 32(6): 12–21
|
|
Wang Zhanchang, Lin Yiming, Feng Danqin, et al. 2009. A new atisane-type diterpene from the bark of the mangrove plant Excoecaria agallocha. Molecules, 14(1): 414–422. doi: 10.3390/molecules14010414
|
|
Watson J. 1988. The cheirolepidiaceae. In: Beck C B, ed. Origin and Evolution of Gymnosperms. New York, NY, USA: Columbia University Press, 382–447
|
|
Zahirovic S, Seton M, Müller R D. 2014. The Cretaceous and Cenozoic tectonic evolution of Southeast Asia. Solid Earth, 5(1): 227–273. doi: 10.5194/se-5-227-2014
|
|
Zhang Gongcheng, Feng Congjun, Yao Xingzong, et al. 2021a. Petroleum geology in deepwater settings in a passive continental margin of a marginal sea: a case study from the South China Sea. Acta Geologica Sinica (English Edition), 95(1): 1–20. doi: 10.1111/1755-6724.14621
|
|
Zhang Gongcheng, Jia Qingjun, Wang Wanyin, et al. 2018. On tectonic framework and evolution of the South China Sea. Chinese Journal of Geophysics, 61(10): 4194–4215
|
|
Zhang Gongcheng, Wang Dongdong, Lan Lei, et al. 2021b. The geological characteristics of the large- and medium-sized gas fields in the South China Sea. Acta Oceanologica Sinica, 40(2): 1–12. doi: 10.1007/s13131-021-1754-x
|
|
Zhong Xiaoyong, Yuan Qin, Qin Zhanjie, et al. 2012. The sporo-pollen analyses and ore-forming age of Nong Bok formation in Khammouane, Laos. Acta Geoscientica Sinica, 33(3): 323–330
|
|
Zhou Hantao, Lin Peng. 2001. Analysis on genetic diversity of mangrove species of Sonneratia and relationship to plant introduction. Acta Oceanologica Sinica, 20(3): 427–434
|


Figures(8) / Tables(1)

Supported by:
Beijing Renhe Information Technology Co. Ltd

Lei Lan, Youchuan Li, Zhigang Zhao, Shuchun Yang, Qing Lin, Weilai Zhang. The influence of organic sources and environments on source rock deposition during the periods of Cretaceous–Eocene and Oligocene–Miocene, northern Kalimantan[J]. Acta Oceanologica Sinica, 2023, 42(3): 54-64. doi: 10.1007/s13131-022-2080-7

| Sample ID | Period/series | Facies | P1 | P2 | P3 | P4 | P5 | P6 | P7 | P8 | P9 | P10 | P11 | P12 | P13 | P14 | P15 |
| K1 | C.–E. | B | –24.0 | 1.1 | 468 | 2.6 | 43 | 0.83 | 0.08 | 0.11 | 0.98 | 0.10 | 0.12 | 0.11 | 0.40 | 6.88 | 1.95 |
| K2 | C.–E. | B | –23.9 | 0.6 | 436 | 2.5 | 22 | N.A. | 0.07 | 0.72 | 1.04 | 0.11 | 0.12 | 0.13 | 0.58 | 8.79 | 2.06 |
| K3 | C.–E. | B | –23.8 | 0.5 | 449 | 1.7 | 31 | 0.83 | 0.09 | 0.75 | 0.85 | 0.14 | 0.24 | 0.10 | 0.29 | 13.00 | 1.95 |
| K4 | C.–E. | B | N.A. | 0.7 | 484 | 1.3 | 56 | N.A. | 0.07 | 0.14 | 0.39 | 0.27 | 0.46 | 0.16 | 0.88 | 6.09 | 1.96 |
| K5 | C.–E. | B | –22.0 | 0.1 | 593 | 2.1 | 3 | 1.47 | 0.06 | 0.06 | 0.25 | 0.41 | 0.80 | 0.14 | 0.23 | 6.87 | 1.74 |
| K6 | C.–E. | B | –23.5 | 0.1 | 588 | 2.7 | 5 | 1.12 | 0.08 | 0.00 | 0.39 | 0.40 | 0.71 | 0.15 | 0.31 | 5.99 | 1.86 |
| K7 | C.–E. | B | –24.6 | 0.1 | 369 | 0.9 | 8 | 2.27 | 0.07 | 0.04 | 0.34 | 0.47 | 0.74 | 0.15 | 0.25 | 5.27 | 1.64 |
| K8 | C.–E. | B | –24.8 | 0.0 | 477 | 0.8 | 4 | 3.61 | 0.05 | 0.21 | 0.36 | 0.43 | 0.82 | 0.15 | 0.34 | 5.05 | 1.74 |
| K9 | C.–E. | B | –26.2 | 0.1 | 597 | 1.5 | 5 | 2.42 | 0.07 | 0.04 | 0.25 | 0.51 | 0.77 | 0.15 | 0.19 | 4.74 | 1.52 |
| K10 | C.–E. | B | –26.7 | 0.1 | 596 | 2.6 | 4 | 2.26 | 0.07 | 0.03 | 0.36 | 0.43 | 0.73 | 0.14 | 0.22 | 4.71 | 1.77 |
| K11 | C.–E. | B | –27.3 | 0.1 | 598 | 3.0 | 4 | 2.57 | 0.07 | 0.05 | 0.38 | 0.33 | 0.57 | 0.17 | 0.38 | 5.15 | 1.40 |
| K12 | C.–E. | B | –24.6 | 0.2 | 448 | 0.6 | 30 | N.A. | 0.02 | 0.07 | 0.56 | 0.52 | 0.73 | 0.06 | 0.35 | 1.57 | 0.99 |
| Q1 | C.–E. | B | –27.2 | 0.0 | 576 | 0.8 | 4 | 1.50 | 0.07 | 0.01 | 0.36 | 0.60 | 1.11 | 0.12 | 0.34 | 4.52 | 1.54 |
| Q2 | C.–E. | B | –27.8 | 0.0 | 477 | 0.6 | 5 | 3.64 | 0.07 | 0.02 | 0.47 | 0.56 | 0.91 | 0.14 | 0.25 | 3.81 | 1.64 |
| Q3 | C.–E. | B | –27.9 | 0.3 | 461 | 0.4 | 77 | N.A. | 0.11 | 0.07 | 0.78 | 0.36 | 0.43 | 0.15 | 0.48 | 4.95 | 1.44 |
| Q4 | C.–E. | B | –26.8 | 0.1 | 597 | 0.5 | 10 | 2.66 | 0.06 | 0.02 | 0.54 | N.D. | N.D. | 0.12 | 0.44 | 4.65 | 1.54 |
| Q5 | O.–M. | B | –26.8 | 0.6 | 457 | 0.9 | 67 | N.A. | 0.46 | 8.72 | 2.53 | 1.48 | 0.57 | 0.11 | 0.48 | 4.40 | 0.28 |
| Q6 | O.–M. | B | –26.5 | 0.3 | 439 | 0.4 | 73 | N.A. | 0.16 | 2.12 | 2.43 | 2.53 | 0.98 | 0.10 | 0.68 | 0.71 | 0.76 |
| Q7 | O.–M. | B | –27.0 | 0.3 | 444 | 0.3 | 84 | 0.81 | 0.21 | 1.19 | 1.97 | 1.69 | 0.95 | 0.11 | 0.49 | 0.52 | 1.33 |
| Q8 | O.–M. | B | –28.4 | 0.3 | 415 | 1.0 | 29 | 0.79 | 0.66 | 1.21 | 1.27 | 1.29 | 1.12 | 0.05 | 0.35 | 2.68 | 0.77 |
| K13 | O.–M. | D-N | –26.0 | 1.0 | 527 | 3.2 | 33 | 0.95 | 0.13 | 0.17 | 0.12 | 0.37 | 0.76 | 0.11 | 0.26 | 2.40 | 1.51 |
| K14 | O.–M. | D-N | N.A. | 2.6 | 443 | 2.7 | 98 | N.A. | 0.21 | 3.35 | 3.10 | 2.11 | 0.62 | 0.05 | 0.39 | 1.44 | 0.47 |
| Q9 | O.–M. | D-N | –27.8 | 4.5 | 448 | 2.9 | 154 | 0.47 | 1.63 | 3.69 | 3.11 | 2.06 | 0.61 | 0.10 | 1.39 | 0.63 | 0.58 |
| Q10 | O.–M. | D-N | –27.0 | 0.4 | 433 | 0.4 | 120 | N.A. | 0.26 | 1.12 | 3.13 | 3.86 | 1.10 | 0.11 | 0.73 | 0.49 | 0.27 |
| Q11 | O.–M. | D-N | –26.5 | 0.2 | 435 | 0.3 | 70 | 1.66 | 0.08 | 1.36 | 1.39 | 1.34 | 0.89 | 0.07 | 0.68 | 0.94 | 0.78 |
| Q12 | O.–M. | D-N | –27.2 | 0.7 | 440 | 0.6 | 107 | 0.39 | 0.19 | 1.20 | 2.49 | 2.21 | 0.89 | 0.08 | 1.01 | 0.32 | 0.62 |
| Q13 | O.–M. | D-N | –27.5 | 0.8 | 442 | 0.7 | 109 | 0.49 | 0.19 | 1.99 | 2.65 | 2.12 | 0.87 | 0.07 | 0.75 | 0.36 | 0.62 |
| Q14 | O.–M. | D-N | –27.1 | 0.5 | 451 | 0.5 | 99 | 1.06 | 0.41 | 1.43 | 2.56 | 2.07 | 0.79 | 0.08 | 1.40 | 0.30 | 0.57 |
| Q15 | O.-M. | D-N | –27.1 | 1.2 | 411 | 1.7 | 75 | 0.65 | 4.39 | 1.21 | 11.98 | N.D. | 0.36 | 0.06 | 0.69 | 0.82 | 0.34 |
| Q16 | O.–M. | D-N | –28.3 | 1.0 | 441 | 0.7 | 133 | N.A. | 0.31 | 0.98 | 2.97 | 3.10 | 1.28 | 0.11 | 0.48 | 0.67 | 0.48 |
| Q17 | O.–M. | C | –27.4 | 29.8 | 445 | 18.7 | 159 | 0.61 | 0.14 | 0.11 | 4.55 | 1.09 | 0.20 | 0.07 | 3.47 | 0.39 | 0.77 |
| Q18 | O.–M. | C | –28.0 | 12.3 | 455 | 9.2 | 134 | 0.78 | 0.76 | 36.64 | 3.45 | 1.60 | 0.53 | 0.03 | 1.12 | 1.78 | 1.42 |
| Q19 | O.–M. | C | –27.2 | 145.7 | 441 | 53.3 | 273 | 0.71 | 0.13 | 0.80 | 3.66 | 3.14 | 1.35 | 0.05 | 3.10 | 0.09 | 0.89 |
| Q20 | O.–M. | C | –26.7 | 29.4 | 433 | 14.9 | 197 | 0.63 | 0.20 | 2.26 | 3.37 | 4.09 | 1.86 | 0.08 | 1.42 | 0.09 | 1.02 |
| Q21 | O.–M. | C | –27.9 | 31.9 | 426 | 21.1 | 152 | N.A. | 1.27 | 0.57 | 3.60 | 0.44 | 1.24 | 0.11 | 0.58 | 0.59 | 0.00 |
| Q22 | O.–M. | C | –26.8 | 1.8 | 412 | 7.3 | 25 | 0.40 | 2.03 | 0.80 | 2.54 | 2.82 | 1.68 | 0.04 | 0.16 | 3.59 | 0.27 |
| Note: C.–E.=Cretaceous–Eocene; O.–M.=Oligocene–Miocene; B=bathyal; N-D=delta-neritic; C=coaly; N.A.=not analyzed; N.D.=no data; P1=$ \text{δ}^{13} $Ckerogen; P2= S2, mg/g; P3=Tmax, ℃; P4=total organic carbon (TOC), %; P5=HI=hydrocarbon index, mg/g; P6=Ro, %; P7=Oleanane/C30 $\alpha \beta $ hopane; P8=bicadinane T/C30 $\alpha \beta $ hopane (m/z 412); P9=Pr/Ph; P10=Pr/nC17; P11=Ph/nC18; P12=Gam/C30 $\alpha \beta $ hopane; P13=(C19+C20)/(C23+C24) tricyclic terpanes; P14=(C19−C29) tricyclic terpanes/C30 $\alpha \beta $ hopane; P15=C27/C29 $\alpha \alpha \alpha $ 20R steranes. | |||||||||||||||||
DownLoad:
CSV
 DownLoad:
DownLoad:

 DownLoad:
DownLoad:






