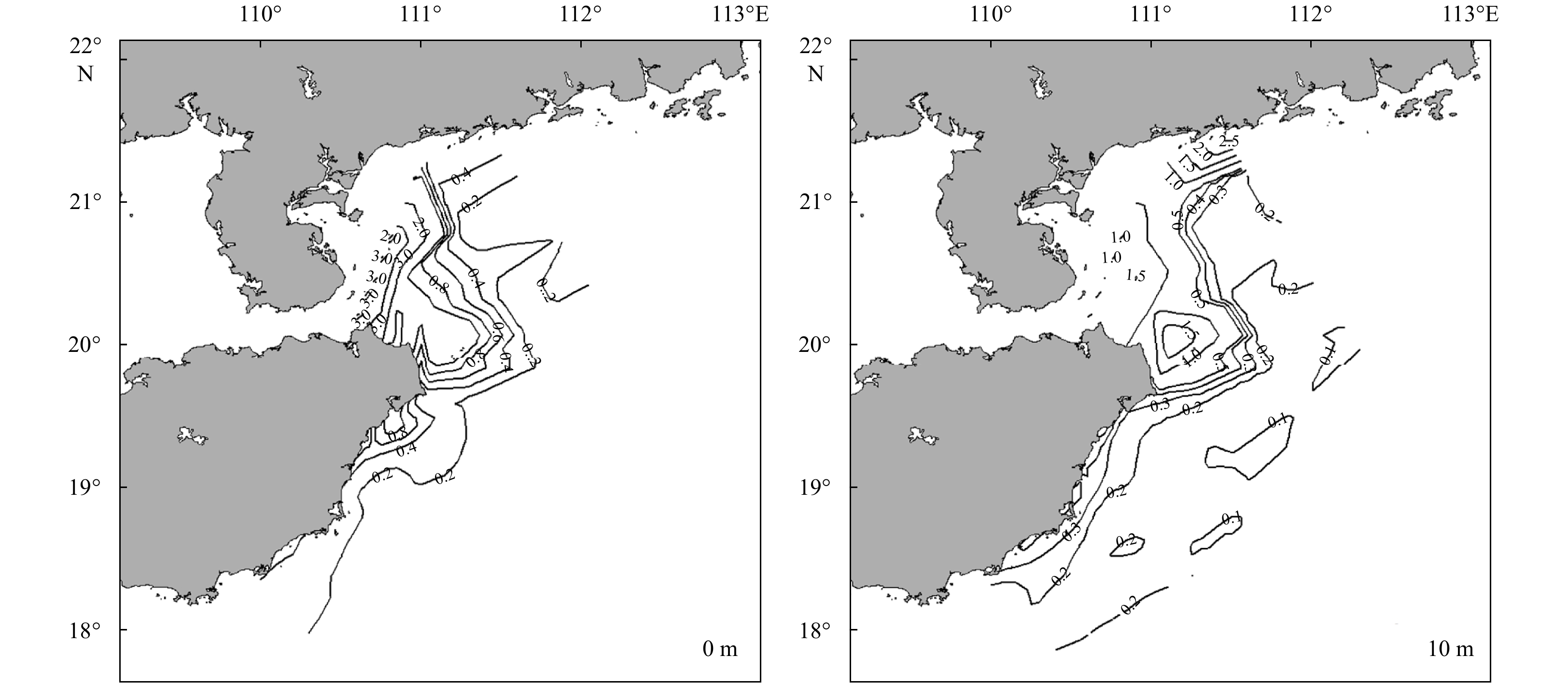
| Citation: | Sumin Liu, Bo Hong, Guifen Wang, Weiqiang Wang, Qiang Xie, Zekai Ni, Liu Yu, Huichang Jiang, Tong Long, Hongzhou Xu. Physical structure and phytoplankton community off the eastern Hainan coast during summer 2015[J]. Acta Oceanologica Sinica, 2020, 39(11): 103-114. doi: 10.1007/s13131-020-1668-z

|

| [1] |
Anglès S, Jordi A, Henrichs D W, et al. 2019. Influence of coastal upwelling and river discharge on the phytoplankton community composition in the northwestern Gulf of Mexico. Progress in Oceanography, 173: 26–36. doi: 10.1016/j.pocean.2019.02.001
|
| [2] |
Benson S R, Croll D A, Marinovic B B, et al. 2002. Changes in the cetacean assemblage of a coastal upwelling ecosystem during El Niño 1997–98 and La Niña 1999. Progress in Oceanography, 54(1–4): 279–291. doi: 10.1016/S0079-6611(02)00054-X
|
| [3] |
Burger J M, Moloney C L, Walker D R, et al. 2020. Drivers of short-term variability in phytoplankton production in an embayment of the southern Benguela upwelling system. Journal of Marine Systems, 208: 103341. doi: 10.1016/j.jmarsys.2020.103341
|
| [4] |
Chen Fajin, Zeng Zhen, Meng Yafei, et al. 2016. Diel variation of nutrients and chlorophyll a concentration in the Qiongdong sea region during the summer of 2013. Haiyang Xuebao (in Chinese), 38(4): 76–83
|
| [5] |
Chen Jixin, Huang Bangqin, Liu Yuan, et al. 2006. Phytoplankton community structure in the transects across East China Sea and northern South China Sea determined by analysis of HPLC photosynthetic pigment signatures. Advances in Earth Science, 21(7): 738–746
|
| [6] |
Deng Song, Zhong Huangliang, Wang Mingwen, et al. 1995. On relation between upwelling off Qionghai and fishery. Journal of Oceanography in Taiwan Strait (in Chinese), 14(1): 51–56
|
| [7] |
Du Xiuning, Peterson W T. 2018. Phytoplankton community structure in 2011–2013 compared to the extratropical warming event of 2014–2015. Geophysical Research Letters, 45(3): 1534–1540. doi: 10.1002/2017GL076199
|
| [8] |
Emeis K, Eggert A, Flohr A, et al. 2018. Biogeochemical processes and turnover rates in the northern Benguela upwelling system. Journal of Marine Systems, 188: 63–80. doi: 10.1016/j.jmarsys.2017.10.001
|
| [9] |
Eppley R W, Thomas W H. 1969. Comparison of half-saturation constants for growth and nitrate uptake of marine phytoplankton. Journal of Phycology, 5(4): 375–379. doi: 10.1111/j.1529-8817.1969.tb02628.x
|
| [10] |
Feng Yuting, Zhao Hui, Shi Yuzhen. 2019. The concentration of nutrients and chlorophyll a in the offshore of Leizhou Peninsula in autumn spatial distribution and their relationship. Journal of Guangdong Ocean University (in Chinese), 39(2): 75–82
|
| [11] |
Ferreira A, Sa C, Silva N, et al. 2020. Phytoplankton response to nutrient pulses in an upwelling system assessed through a microcosm experiment (Algarrobo Bay, Chile). Ocean and Coastal Management, 190: 105167. doi: 10.1016/j.ocecoaman.2020.105167
|
| [12] |
Gan Jianping, Cheung A, Guo Xiaogang, et al. 2009. Intensified upwelling over a widened shelf in the northeastern South China Sea. Journal of Geophysical Research: Oceans, 114(C9): C09019. doi: 10.1029/2007JC004660
|
| [13] |
Guo Fei, Shi Maochong, Xia Zongwan. 1998. Two-demension diagnose model to calculate upwelling on offshore of the east coast of Hainan Island. Haiyang Xuebao (in Chinese), 20(6): 109–116
|
| [14] |
Hallegraeff G M. 1981. Seasonal study of phytoplankton pigments and species at a coastal station off Sydney: Importance of diatoms and the nanoplankton. Marine Biology, 61(2−3): 107–118. doi: 10.1007/BF00386650
|
| [15] |
Han Wuying, Wang Mingbiao, Ma Kemei. 1990. On the lowest surface water temperature area of China Sea in summer-the upwelling along the east coast of Hainan island. Oceanologia et Limnologia Sinica (in Chinese), 21(3): 267–275
|
| [16] |
Hirata T, Hardman-mountford N J, Brewin R J W, et al. 2011. Synoptic relationships between surface chlorophyll-a and diagnostic pigments specific to phytoplankton functional types. Biogeosciences, 8(2): 311–327. doi: 10.5194/bg-8-311-2011
|
| [17] |
Hong Qiming, Li Li. 1991. A study of upwelling over continental shelf off eastern Guangdong. Journal of Oceanography in Taiwan Strait (in Chinese), 10(3): 271–277
|
| [18] |
Huang Bangqin, Hu Jun, Xu Hongzhou, et al. 2010. Phytoplankton community at warm eddies in the northern South China Sea in winter 2003/2004. Deep Sea Research Part II: Topical Studies in Oceanography, 57(19−20): 1792–1798. doi: 10.1016/j.dsr2.2010.04.005
|
| [19] |
Jing Zhiyou, Qi Yiquan, Du Yan. 2011. Upwelling in the continental shelf of northern South China Sea associated with 1997–98 El Niño. Journal of Geophysical Research: Oceans, 116(C2): C02033. doi: 10.1029/2010JC006598
|
| [20] |
Jing Zhiyou, Qi Yiquan, Du Yan, et al. 2015. Summer upwelling and thermal fronts in the northwestern South China Sea: Observational analysis of two mesoscale mapping surveys. Journal of Geophysical Research: Oceans, 120(3): 1993–2006. doi: 10.1002/2014JC010601
|
| [21] |
Jing Zhiyou, Qi Yiquan, Hua Zulin, et al. 2009. Numerical study on the summer upwelling system in the northern continental shelf of the South China Sea. Continental Shelf Research, 29(2): 467–478. doi: 10.1016/j.csr.2008.11.008
|
| [22] |
Lü Xingang, Qiao Fangli, Wang Guansuo, et al. 2008. Upwelling off the west coast of Hainan Island in summer: Its detection and mechanisms. Geophysical Research Letters, 35(2): L02604. doi: 10.1029/2007GL032440
|
| [23] |
Lü Xingang, Qiao Fangli, Xia Changshui, et al. 2010. Upwelling and surface cold patches in the Yellow Sea in summer: Effects of tidal mixing on the vertical circulation. Continental Shelf Research, 30(6): 620–632. doi: 10.1016/j.csr.2009.09.002
|
| [24] |
Lambert C D, Bianchi T S, Santschi P H. 1999. Cross-shelf changes in phytoplankton community composition in the Gulf of Mexico (Texas shelf/slope): The use of plant pigments as biomarkers. Continental Shelf Research, 19(1): 1–21. doi: 10.1016/S0278-4343(98)00075-2
|
| [25] |
Lehmann A, Myrberg K. 2008. Upwelling in the Baltic Sea—A review. Journal of Marine Systems, 74(S1): S3–S12
|
| [26] |
Li Li. 1993. Summer upwelling system over the northern continental shelf of the South China Sea—Physical description. In: Su J, Chuang W S, Hsurh R Y, eds. Proceedings of the Symposium on the Physical and Chemical Oceanography of the China Seas. Beijing: China Ocean Press, 58–68
|
| [27] |
Li Li, Liu Jie, He Juan, et al. 2014. Factors affecting the abundance and community structure of the phytoplankton in northern South China Sea in the summer of 2008: A biomarker study. Chinese Science Bulletin, 59(10): 981–991. doi: 10.1007/s11434-013-0106-4
|
| [28] |
Li Yineng, Peng Shiqiu, Yang Wei, et al. 2012. Numerical simulation of the structure and variation of upwelling off the east coast of Hainan Island using QuikSCAT winds. Chinese Journal of Oceanology and Limnology, 30(6): 1068–1081. doi: 10.1007/s00343-012-1275-8
|
| [29] |
Li Kaizhi, Yin Jianqiang, Huang Liangmin, et al. 2010. Monsoon-forced distribution and assemblages of appendicularians in the northwestern coastal waters of South China Sea. Estuarine, Coastal and Shelf Science, 89(2): 145–153. doi: 10.1016/j.ecss.2010.06.001
|
| [30] |
Li Kaizhi, Yin Jianqiang, Huang Liangmin, et al. 2011. Distribution and abundance of thaliaceans in the northwest continental shelf of South China Sea, with response to environmental factors driven by monsoon. Continental Shelf Research, 31(9): 979–989. doi: 10.1016/j.csr.2011.03.004
|
| [31] |
Li Q P, Zhou Weiwen, Chen Yinchao, et al. 2018. Phytoplankton response to a plume front in the northern South China Sea. Biogeosciences, 15(8): 2551–2563. doi: 10.5194/bg-15-2551-2018
|
| [32] |
Lin Peigen, Hu Jianyu, Zheng Quanan, et al. 2016. Observation of summertime upwelling off the eastern and northeastern coasts of Hainan Island, China. Ocean Dynamics, 66(3): 387–399. doi: 10.1007/s10236-016-0934-2
|
| [33] |
Liu Yi, Peng Zicheng, Wei Gangjian, et al. 2009. Variation of summer coastal upwelling at northern South China Sea during the last 100 years. Geochimica (in Chinese), 38(4): 317–322
|
| [34] |
Örnólfsdóttir E B, Lumsden S E, Pinckney J L. 2004. Phytoplankton community growth-rate response to nutrient pulses in a shallow turbid estuary, Galveston Bay, Texas. Journal of Plankton Research, 26(3): 325–339. doi: 10.1093/plankt/fbh035
|
| [35] |
Pauly D, Christensen V. 1995. Primary production required to sustain global fisheries. Nature, 374(6519): 255–257. doi: 10.1038/374255a0
|
| [36] |
Peng Xin, Ning Xiuren, Cai Yiming, et al. 2006. Review of research on the bottom-up effects of phytoplankton growth. Journal of Marine Sciences (in Chinese), 24(3): 64–75
|
| [37] |
Roy C, Reason C. 2001. ENSO related modulation of coastal upwelling in the eastern Atlantic. Progress in Oceanography, 49(1−4): 245–255. doi: 10.1016/S0079-6611(01)00025-8
|
| [38] |
Sañé E, Valente A, Fatela F, et al. 2019. Assessment of sedimentary pigments and phytoplankton determined by CHEMTAX analysis as biomarkers of unusual upwelling conditions in summer 2014 off the SE coast of Algarve. Journal of Sea Research, 146: 33–45. doi: 10.1016/j.seares.2019.01.007
|
| [39] |
Shu Yeqiang, Wang Dongxiao, Feng Ming, et al. 2018. The contribution of local wind and ocean circulation to the interannual variability in coastal upwelling intensity in the northern South China Sea. Journal of Geophysical Research: Oceans, 123(9): 6766–6778. doi: 10.1029/2018JC014223
|
| [40] |
Sieburth J M, Smetacek V, Lenz J. 1978. Pelagic ecosystem structure: Heterotrophic compartments of the plankton and their relationship to plankton size fractions. Limnology and Oceanography, 23(6): 1256–1263. doi: 10.4319/lo.1978.23.6.1256
|
| [41] |
Silkin V A, Pautova L A, Giordano M, et al. 2019. Drivers of phytoplankton blooms in the northeastern Black Sea. Marine Pollution Bulletin, 138: 274–284. doi: 10.1016/j.marpolbul.2018.11.042
|
| [42] |
Smith R L. 1995. The physical processes of coastal ocean upwelling systems. In: Summerhayes C P, Emeis K C, Angel M V, et al, eds. Upwelling in the Ocean: Modern Processes and Ancient Records. Chichester: John Wiley and Sons, 18: 39–64
|
| [43] |
Sobarzo M, Bravo L, Donoso D, et al. 2007. Coastal upwelling and seasonal cycles that influence the water column over the continental shelf off central Chile. Progress in Oceanography, 75(3): 363–382. doi: 10.1016/j.pocean.2007.08.022
|
| [44] |
Su Jian, Pohlmann T. 2009. Wind and topography influence on an upwelling system at the eastern Hainan coast. Journal of Geophysical Research: Oceans, 114(C6): C06017. doi: 10.1029/2008JC005018
|
| [45] |
Su Jian, Wang Jun, Pohlmann T, et al. 2011. The influence of meteorological variation on the upwelling system off eastern Hainan during summer 2007–2008. Ocean Dynamics, 61(6): 717–730. doi: 10.1007/s10236-011-0404-9
|
| [46] |
Su Jian, Xu Mingquan, Pohlmann T, et al. 2013. A western boundary upwelling system response to recent climate variation (1960–2006). Continental Shelf Research, 57: 3–9. doi: 10.1016/j.csr.2012.05.010
|
| [47] |
Tilman D, Kiesling R, Sterner R, et al. 1986. Green, blue-green and diatom algae: taxonomic differences in competitive ability for phosphorus, silicon and nitrogen. Archiv Fur Hydrobiologie, 106(4): 473–485
|
| [48] |
Tsuchiya K, Kuwahara V S, Yoshiki T M, et al. 2013. Phytoplankton community response and succession in relation to typhoon passages in the coastal waters of Japan. Journal of Plankton Research, 36(2): 424–438
|
| [49] |
van Heukelem L, Thomas C S. 2001. Computer-assisted high-performance liquid chromatography method development with applications to the isolation and analysis of phytoplankton pigments. Journal of Chromatography A, 910(1): 31–49. doi: 10.1016/S0378-4347(00)00603-4
|
| [50] |
Wang Mengyin, Hu Qiwei. 2017. Impact of summer upwelling on the fisheries resources in the northern South China Sea based on remote sensing data. Journal of Hainan Tropical Ocean University (in Chinese), 24(2): 22–29
|
| [51] |
Wang Yu, Kang Jianhua, Ye Youyin, et al. 2016. Phytoplankton community and environmental correlates in a coastal upwelling zone along western Taiwan Strait. Journal of Marine Systems, 154: 252–263. doi: 10.1016/j.jmarsys.2015.10.015
|
| [52] |
Wang Dongxiao, Shu Yeqiang, Xue Huijie, et al. 2014. Relative contributions of local wind and topography to the coastal upwelling intensity in the northern South China Sea. Journal of Geophysical Research: Oceans, 119(4): 2550–2567. doi: 10.1002/2013JC009172
|
| [53] |
Wang Daoru, Yang Yi, Wang Jia, et al. 2015. A modeling study of the effects of river runoff, tides, and surface wind-wave mixing on the eastern and western Hainan upwelling systems of the South China Sea, China. Ocean Dynamics, 65(8): 1143–1164. doi: 10.1007/s10236-015-0857-3
|
| [54] |
Wu Risheng, Li Li. 2003. Summarization of study on upwelling system in the South China Sea. Journal of Oceanography in Taiwan Strait (in Chinese), 22(2): 269–277
|
| [55] |
Xie Lingling, He Chaofeng, Li Mingming, et al. 2017. Response of sea surface temperature to Typhoon passages over the upwelling zone east of Hainan Island. Advances in Marine Science (in Chinese), 35(1): 8–19
|
| [56] |
Xie Shangping, Xie Qiang, Wang Dongxiao, et al. 2003. Summer upwelling in the South China Sea and its role in regional climate variations. Journal of Geophysical Research: Oceans, 108(C8): 3261. doi: 10.1029/2003JC001867
|
| [57] |
Xie Lingling, Zhang Shuwen, Zhao Hui. 2012. Overview of studies on Qiongdong upwelling. Journal of Tropical Oceanography (in Chinese), 31(4): 35–41
|
| [58] |
Xie Lingling, Zong Xiaolong, Yi Xiaofei, et al. 2016. The interannual variation and long-term trend of qiongdong upwelling. Oceanologia et Limnologia Sinica (in Chinese), 47(1): 43–51
|
| [59] |
Xu Jindian, Cai Shangzhan, Xuan Lili, et al. 2013. Study on coastal upwelling in eastern Hainan Island and western Guangdong in summer, 2006. Acta Oceanologica Sinica (in Chinese), 35(4): 11–18
|
| [60] |
Xue Yan, Kumar A. 2017. Evolution of the 2015/16 El Niño and historical perspective since 1979. Science China Earth Science, 60(9): 1572–1588. doi: 10.1007/s11430-016-0106-9
|
| [61] |
Yang Dezhou, Yin Baoshu, Sun Junchuan, et al. 2013. Numerical study on the origins and the forcing mechanism of the phosphate in upwelling areas off the coast of Zhejiang province, China in summer. Journal of Marine Systems, 123–124: 1–18. doi: 10.1016/j.jmarsys.2013.04.002
|
| [62] |
Yin Jianqiang, Huang Liangmin, Li Kaizhi, et al. 2011. Abundance distribution and seasonal variations of Calanus sinicus (Copepoda: Calanoida) in the northwest continental shelf of South China Sea. Continental Shelf Research, 31(14): 1447–1456. doi: 10.1016/j.csr.2011.06.010
|
| [63] |
Zeng Zhen, Chen Fajin, Meng Yafei, et al. 2015. Nutrient structure and its influence on phytoplankton growth in the Qiongdong Sea during winter and summer. Journal of Guangdong Ocean University (in Chinese), 35(3): 70–77
|
| [64] |
Zhai Hongchang, Ning Xiuren, Tang Xuexi, et al. 2011. Phytoplankton pigment patterns and community composition in the northern South China Sea during winter. Chinese Journal of Oceanology and Limnology, 29(2): 233–245. doi: 10.1007/s00343-011-0111-x
|

Figures(10) / Tables(3)

Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: