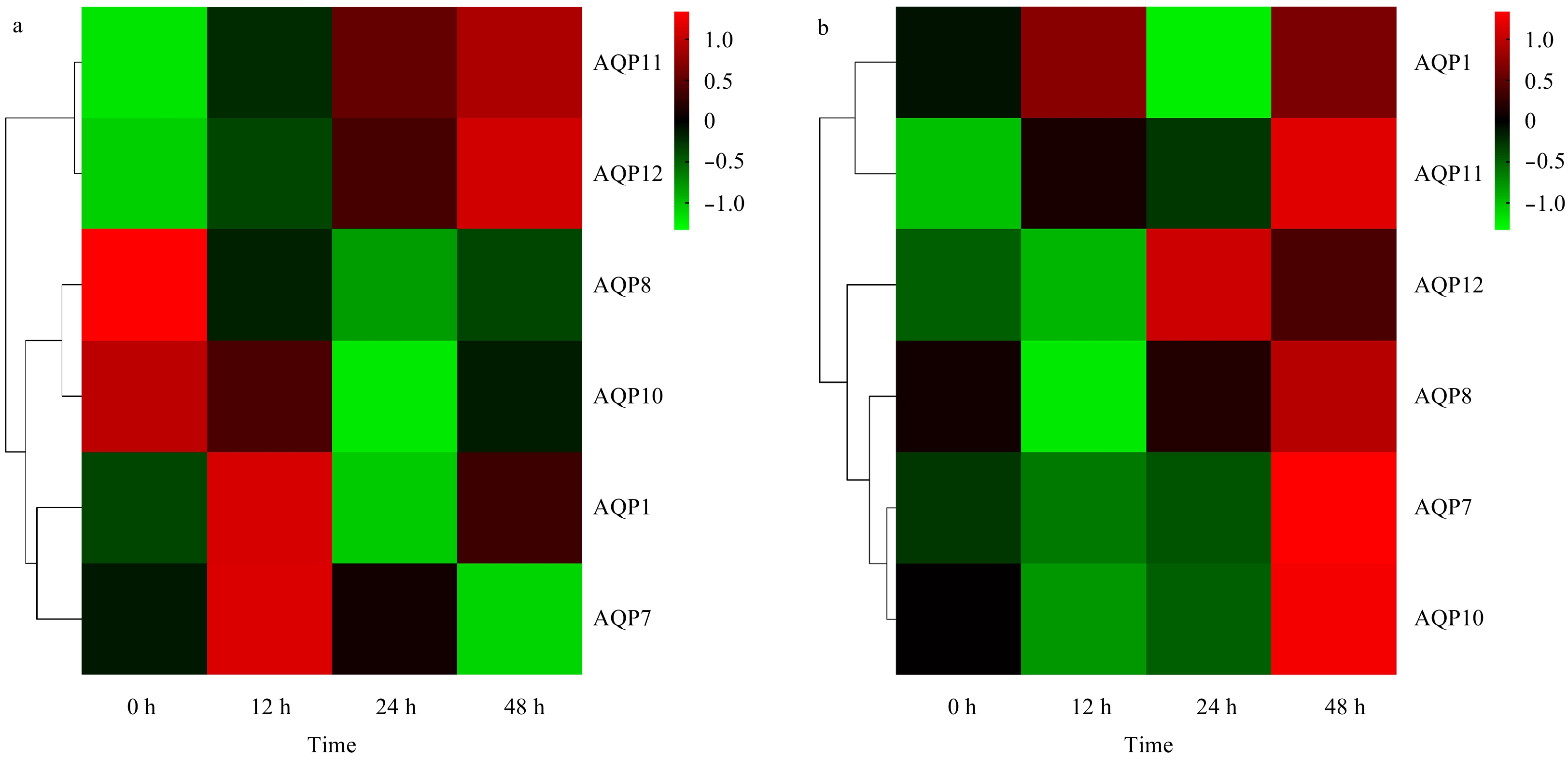
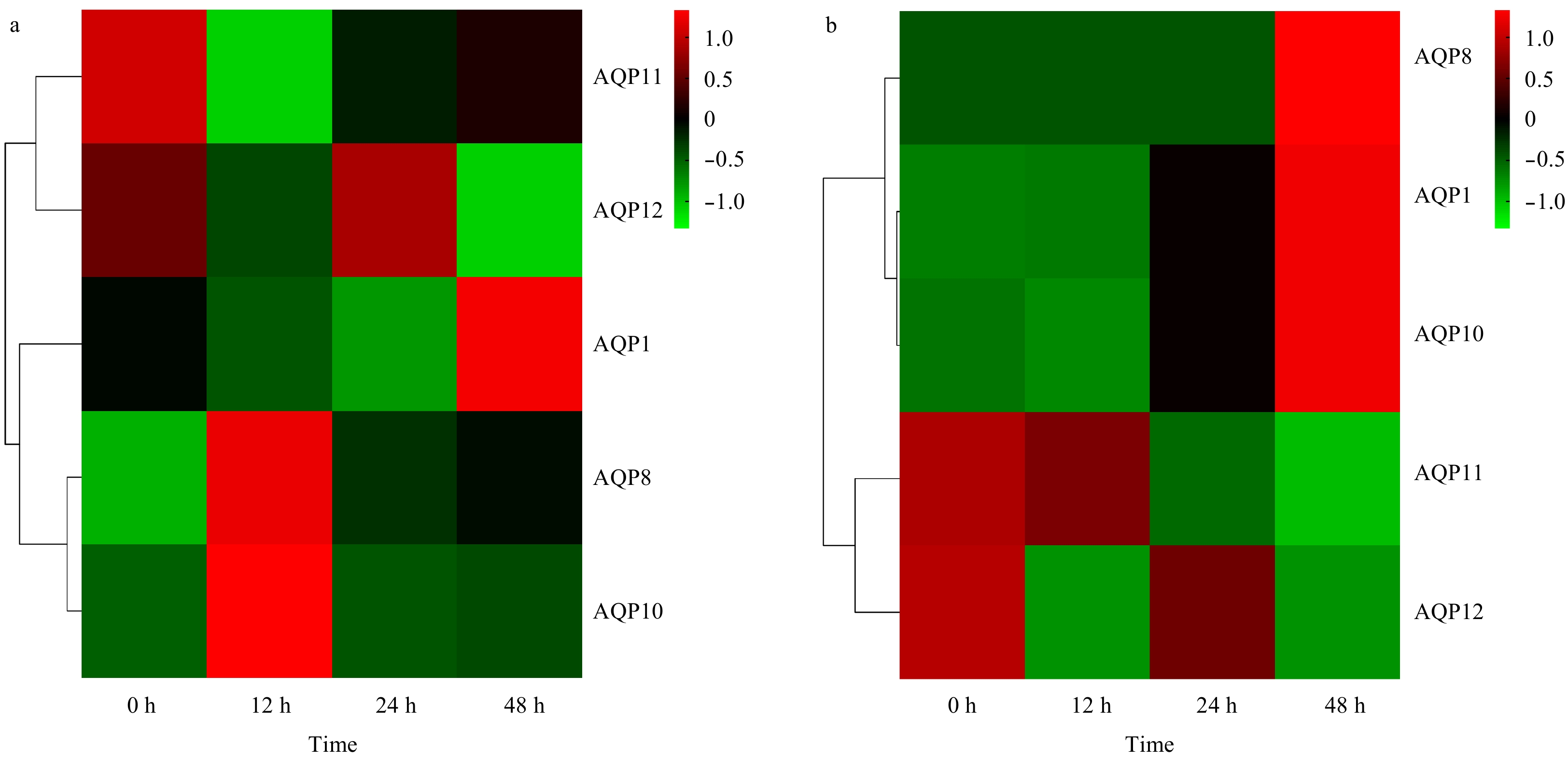
| Citation: | Qian Ma, Xinfu Liu, Ang Li, Shufang Liu, Zhimeng Zhuang. Effects of osmotic stress on the expression profiling of aquaporin genes in the roughskin sculpin (Trachidermus fasciatus)[J]. Acta Oceanologica Sinica, 2020, 39(6): 19-25. doi: 10.1007/s13131-020-1594-0

|

| [1] |
Agre P, King L S, Yasui M, et al. 2002. Aquaporin water channels-from atomic structure to clinical medicine. Journal of Physiology, 542(1): 3–16. doi: 10.1113/jphysiol.2002.020818
|
| [2] |
An K W, Kim N N, Choi C Y. 2008. Cloning and expression of aquaporin 1 and arginine vasotocin receptor mRNA from the black porgy, Acanthopagrus schlegeli: effect of freshwater acclimation. Fish Physiology and Biochemistry, 34(2): 185–194. doi: 10.1007/s10695-007-9175-0
|
| [3] |
Aoki M, Kaneko T, Katoh F, et al. 2003. Intestinal water absorption through aquaporin 1 expressed in the apical membrane of mucosal epithelial cells in seawater-adapted Japanese eel. Journal of Experimental Biology, 206(19): 3495–3505. doi: 10.1242/jeb.00579
|
| [4] |
Cao Jun, Shi Feng. 2019. Comparative analysis of the aquaporin gene family in 12 fish species. Animals, 9(5): 233. doi: 10.3390/ani9050233
|
| [5] |
Cerdà J, Finn R N. 2010. Piscine aquaporins: an overview of recent advances. Journal of Experimental Zoology Part A, 313A(10): 623–650. doi: 10.1002/jez.634
|
| [6] |
Choi Y J, Shin H S, Kim N N, et al. 2013. Expression of aquaporin-3 and -8 mRNAs in the parr and smolt stages of sockeye salmon, Oncorhynchus nerka: effects of cortisol treatment and seawater acclimation. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 165(2): 228–236
|
| [7] |
Cutler C P, Cramb G. 2000. Water transport and aquaporin expression in fish. In: Hohmann S, Nielsen S, eds. Molecular Biology and Physiology of Water and Solute Transport. Boston: Springer, 433–441
|
| [8] |
Cutler C P, Cramb G. 2002. Branchial expression of an aquaporin 3(AQP-3) homologue is downregulated in the European eel Anguilla anguilla following seawater acclimation. Journal of Experimental Biology, 205(17): 2643–2651
|
| [9] |
Cutler C P, Martinez A S, Cramb G. 2007. The role of aquaporin 3 in teleost fish. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 148(1): 82–91
|
| [10] |
Cutler C P, Philips C, Hazon N, et al. 2009. Aquaporin 8 (AQP8) intestinal mRNA expression increases in response to salinity acclimation in yellow and silver European eels (Anguilla anguilla). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 153(2): S78
|
| [11] |
Engelund M B, Madsen S S. 2011. The role of aquaporins in the kidney of euryhaline teleosts. Frontiers in Physiology, 2: 51
|
| [12] |
Engelund M B, Madsen S S. 2015. Tubular localization and expressional dynamics of aquaporins in the kidney of seawater-challenged Atlantic salmon. Journal of Comparative Physiology B, 185(2): 207–223. doi: 10.1007/s00360-014-0878-0
|
| [13] |
Finn R N, Cerdà J. 2011. Aquaporin evolution in fishes. Frontiers in Physiology, 2: 44
|
| [14] |
Giffard-Mena I, Boulo V, Abed C, et al. 2011. Expression and localization of aquaporin 1a in the sea-bass (Dicentrarchus labrax) during Ontogeny. Frontiers in Physiology, 2: 34
|
| [15] |
Giffard-Mena I, Boulo V, Aujoulat F, et al. 2007. Aquaporin molecular characterization in the sea-bass (Dicentrarchus labrax): the effect of salinity on AQP1 and AQP3 expression. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 148(2): 430–444
|
| [16] |
Giffard-Mena I, Lorin-Nebel C, Charmantier G, et al. 2008. Adaptation of the sea-bass (Dicentrarchus labrax) to fresh water: role of aquaporins and Na+/K+-ATPases. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 150(3): 332–338
|
| [17] |
Gonzalez R J. 2012. The physiology of hyper-salinity tolerance in teleost fish: a review. Journal of Comparative Physiology B, 182(3): 321–329. doi: 10.1007/s00360-011-0624-9
|
| [18] |
Gorelick D A, Praetorius J, Tsunenari T, et al. 2006. Aquaporin-11: a channel protein lacking apparent transport function expressed in brain. BMC Biochemistry, 7: 14. doi: 10.1186/1471-2091-7-14
|
| [19] |
Goto A. 1990. Alternative life-history styles of Japanese freshwater sculpins revisited. Environmental Biology of Fishes, 28(1–4): 101–112. doi: 10.1007/BF00751030
|
| [20] |
Hamdi M, Sanchez M A, Beene L C, et al. 2009. Arsenic transport by zebrafish aquaglyceroporins. BMC Molecular Biology, 10: 104. doi: 10.1186/1471-2199-10-104
|
| [21] |
Hara-Chikuma M, Verkman A S. 2006. Physiological roles of glycerol-transporting aquaporins: the aquaglyceroporins. Cellular and Molecular Life Sciences CMLS, 63(12): 1386–1392. doi: 10.1007/s00018-006-6028-4
|
| [22] |
Harper C, Wolf J C. 2009. Morphologic effects of the stress response in fish. Ilar Journal, 50(4): 387–396. doi: 10.1093/ilar.50.4.387
|
| [23] |
Ip Y K, Soh M M L, Chen X L, et al. 2013. Molecular characterization of branchial aquaporin 1aa and effects of seawater acclimation, emersion or ammonia exposure on its mRNA expression in the gills, gut, kidney and skin of the freshwater climbing perch, Anabas testudineus. PLoS One, 8(4): e61163. doi: 10.1371/journal.pone.0061163
|
| [24] |
Itoh T, Rai T, Kuwahara M, et al. 2005. Identification of a novel aquaporin, AQP12, expressed in pancreatic acinar cells. Biochemical and Biophysical Research Communications, 330(3): 832–838. doi: 10.1016/j.bbrc.2005.03.046
|
| [25] |
Jeong S Y, Kim J H, Lee W O, et al. 2014. Salinity changes in the anadromous river pufferfish, Takifugu obscurus, mediate gene regulation. Fish Physiology and Biochemistry, 40(1): 205–219. doi: 10.1007/s10695-013-9837-z
|
| [26] |
Kim Y K, Lee S Y, Kim B S, et al. 2014. Isolation and mRNA expression analysis of aquaporin isoforms in marine medaka Oryzias dancena, a euryhaline teleost. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 171: 1–8
|
| [27] |
Kim Y K, Watanabe S, Kaneko T, et al. 2010. Expression of aquaporins 3, 8 and 10 in the intestines of freshwater- and seawater-acclimated Japanese eels Anguilla japonica. Fisheries Science, 76(4): 695–702. doi: 10.1007/s12562-010-0259-x
|
| [28] |
King L S, Kozono D, Agre P. 2004. From structure to disease: the evolving tale of aquaporin biology. Nature Reviews Molecular Cell Biology, 5(9): 687–698. doi: 10.1038/nrm1469
|
| [29] |
Lema S C, Carvalho P G, Egelston J N, et al. 2018. Dynamics of gene expression responses for ion transport proteins and aquaporins in the gill of a euryhaline pupfish during freshwater and high-salinity acclimation. Physiological and Biochemical Zoology, 91(6): 1148–1171. doi: 10.1086/700432
|
| [30] |
Livak K J, Schmittgen T D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the
|
| [31] |
Ma Qian, Liu Xinfu, Feng Wenrong, et al. 2018. Analyses of the molecular mechanisms associated with salinity adaption of Trachidermus fasciatus through combined iTRAQ-based proteomics and RNA sequencing-based transcriptomics. Progress in Biophysics and Molecular Biology, 136: 40–53. doi: 10.1016/j.pbiomolbio.2018.02.003
|
| [32] |
Madsen S S, Engelund M B, Cutler C P. 2015. Water transport and functional dynamics of aquaporins in osmoregulatory organs of fishes. The Biological Bulletin, 229(1): 70–92. doi: 10.1086/BBLv229n1p70
|
| [33] |
Marshall W S, Grosell M. 2006. Ion transport, osmoregulation, and acid-base balance. The Physiology of Fishes, 3: 177–230
|
| [34] |
Martinez A S, Cutler C P, Wilson G D, et al. 2005. Regulation of expression of two aquaporin homologs in the intestine of the European eel: effects of seawater acclimation and cortisol treatment. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 288(6): R1733–R1743. doi: 10.1152/ajpregu.00747.2004
|
| [35] |
Mobasheri A, Marples D. 2004. Expression of the AQP-1 water channel in normal human tissues: a semiquantitative study using tissue microarray technology. American Journal of Physiology-Cell Physiology, 286(3): C529–C537. doi: 10.1152/ajpcell.00408.2003
|
| [36] |
Nielsen S, Frokiaer J, Marples D, et al. 2002. Aquaporins in the kidney: from molecules to medicine. Physiological Reviews, 82(1): 205–244. doi: 10.1152/physrev.00024.2001
|
| [37] |
Raldúa D, Otero D, Fabra M, et al. 2008. Differential localization and regulation of two aquaporin-1 homologs in the intestinal epithelia of the marine teleost Sparus aurata. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology, 294(3): R993–R1003. doi: 10.1152/ajpregu.00695.2007
|
| [38] |
Santos C R A, Estêvão M D, Fuentes J, et al. 2004. Isolation of a novel aquaglyceroporin from a marine teleost (Sparus auratus): function and tissue distribution. Journal of Experimental Biology, 207(7): 1217–1227. doi: 10.1242/jeb.00867
|
| [39] |
Takata K, Matsuzaki T, Tajika Y. 2004. Aquaporins: water channel proteins of the cell membrane. Progress in Histochemistry and Cytochemistry, 39(1): 1–83. doi: 10.1016/j.proghi.2004.03.001
|
| [40] |
Tingaud-Sequeira A, Calusinska M, Finn R N, et al. 2010. The zebrafish genome encodes the largest vertebrate repertoire of functional aquaporins with dual paralogy and substrate specificities similar to mammals. BMC Evolutionary Biology, 10: 38. doi: 10.1186/1471-2148-10-38
|
| [41] |
Tingaud-Sequeira A, Chauvigné F, Fabra M, et al. 2008. Structural and functional divergence of two fish aquaporin-1 water channels following teleost-specific gene duplication. BMC Evolutionary Biology, 8: 259. doi: 10.1186/1471-2148-8-259
|
| [42] |
Tipsmark C K, Sørensen K J, Madsen S S. 2010. Aquaporin expression dynamics in osmoregulatory tissues of Atlantic salmon during smoltification and seawater acclimation. Journal of Experimental Biology, 213(3): 368–379. doi: 10.1242/jeb.034785
|
| [43] |
Verkman A S, Binder D K, Bloch O, et al. 2006. Three distinct roles of aquaporin-4 in brain function revealed by knockout mice. Biochimica et Biophysica Acta (BBA)—Biomembranes, 1758(8): 1085–1093. doi: 10.1016/j.bbamem.2006.02.018
|
| [44] |
Watanabe S, Kaneko T, Aida K. 2005. Aquaporin-3 expressed in the basolateral membrane of gill chloride cells in Mozambique tilapia Oreochromis mossambicus adapted to freshwater and seawater. Journal of Experimental Biology, 208(14): 2673–2682. doi: 10.1242/jeb.01684
|

Figures(4) / Tables(1)

Supported by:
Beijing Renhe Information Technology Co. Ltd






 DownLoad:
DownLoad: