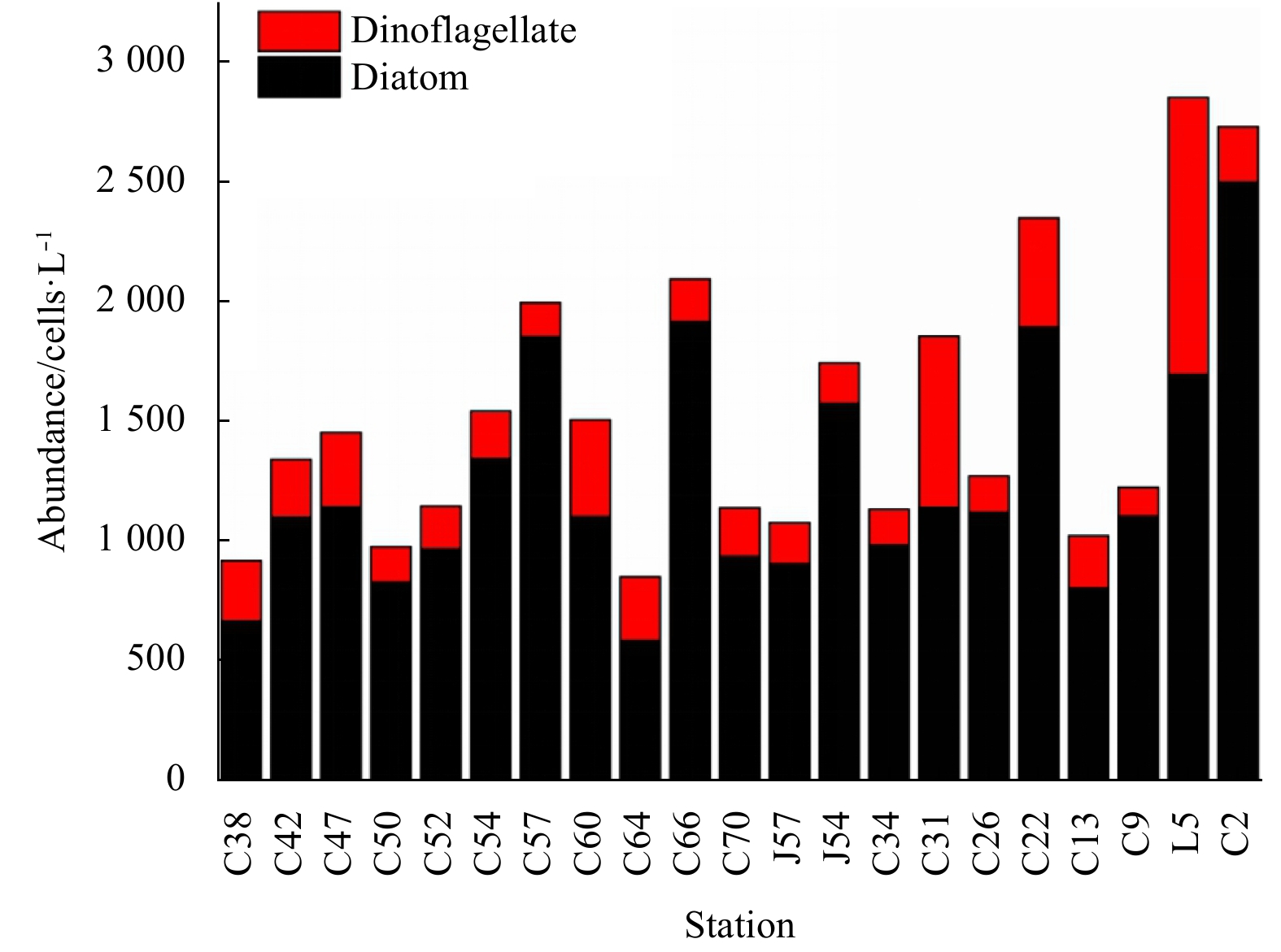
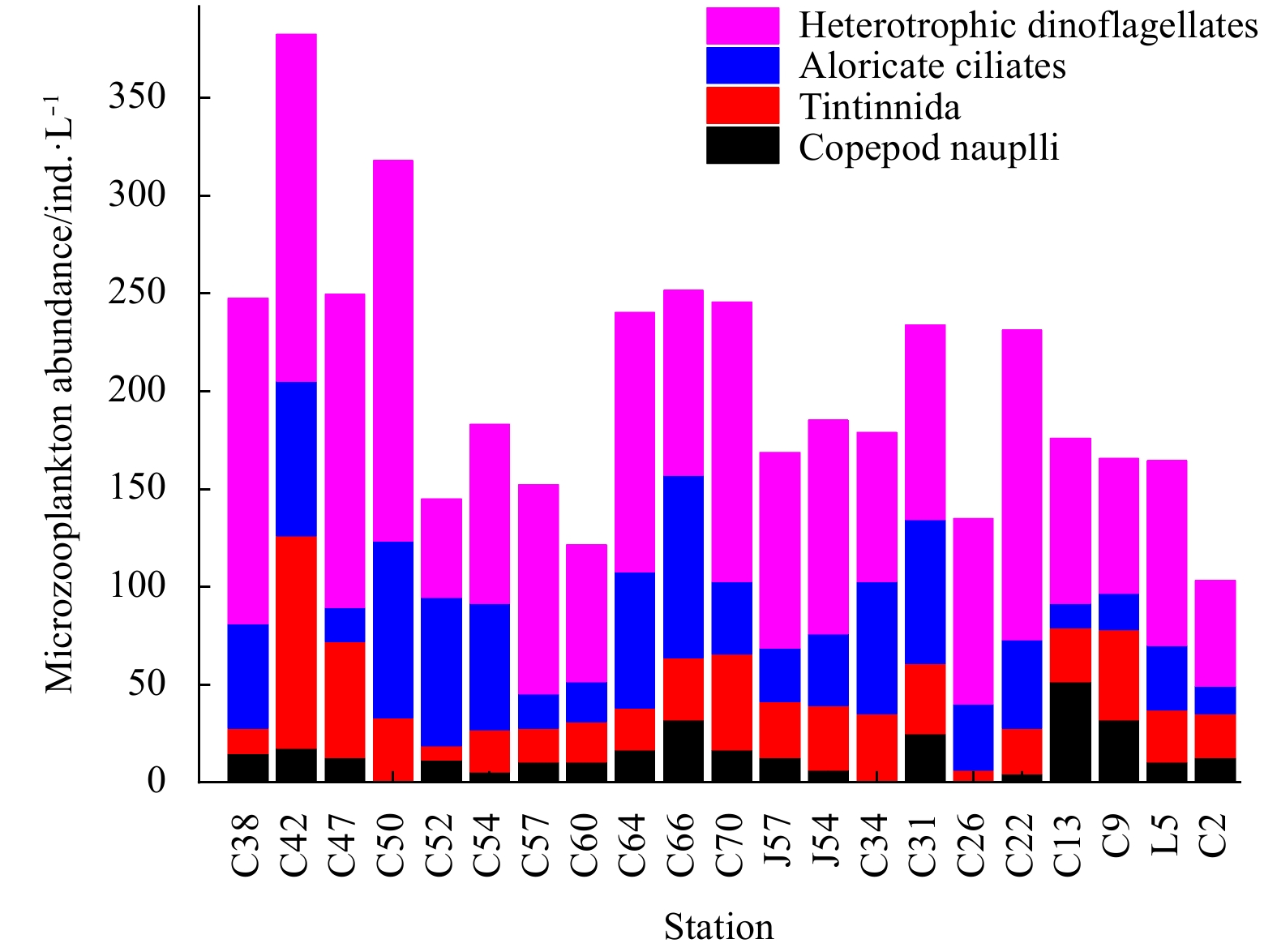
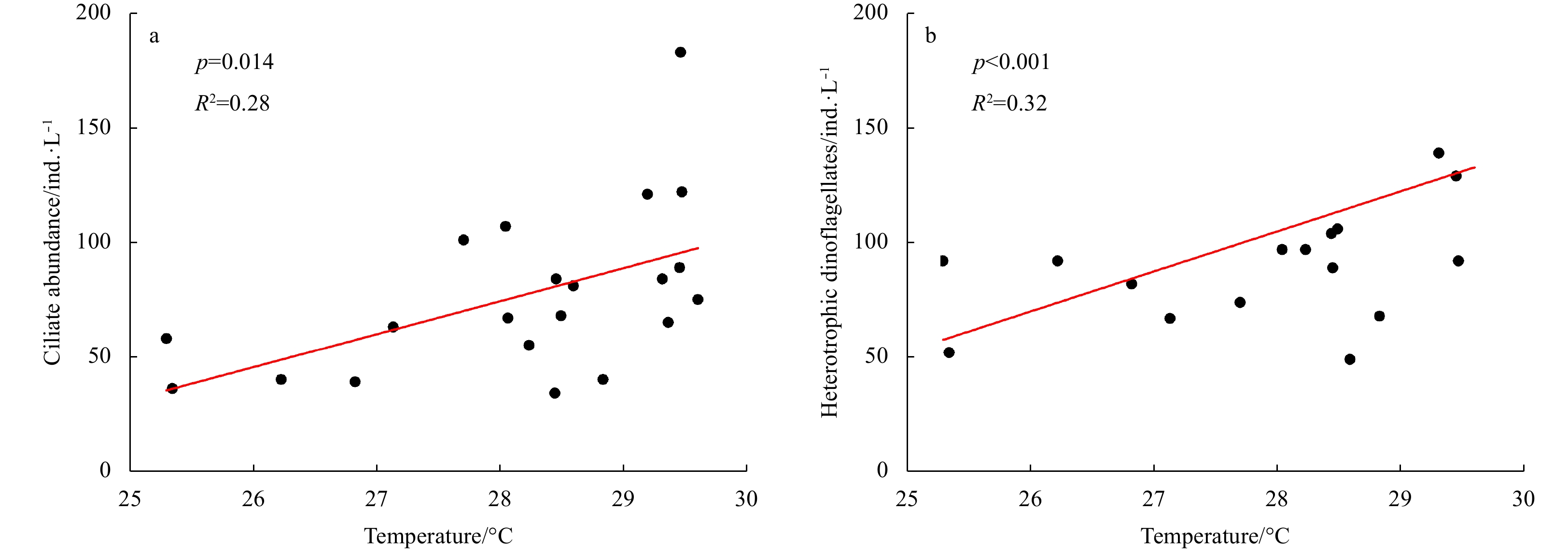
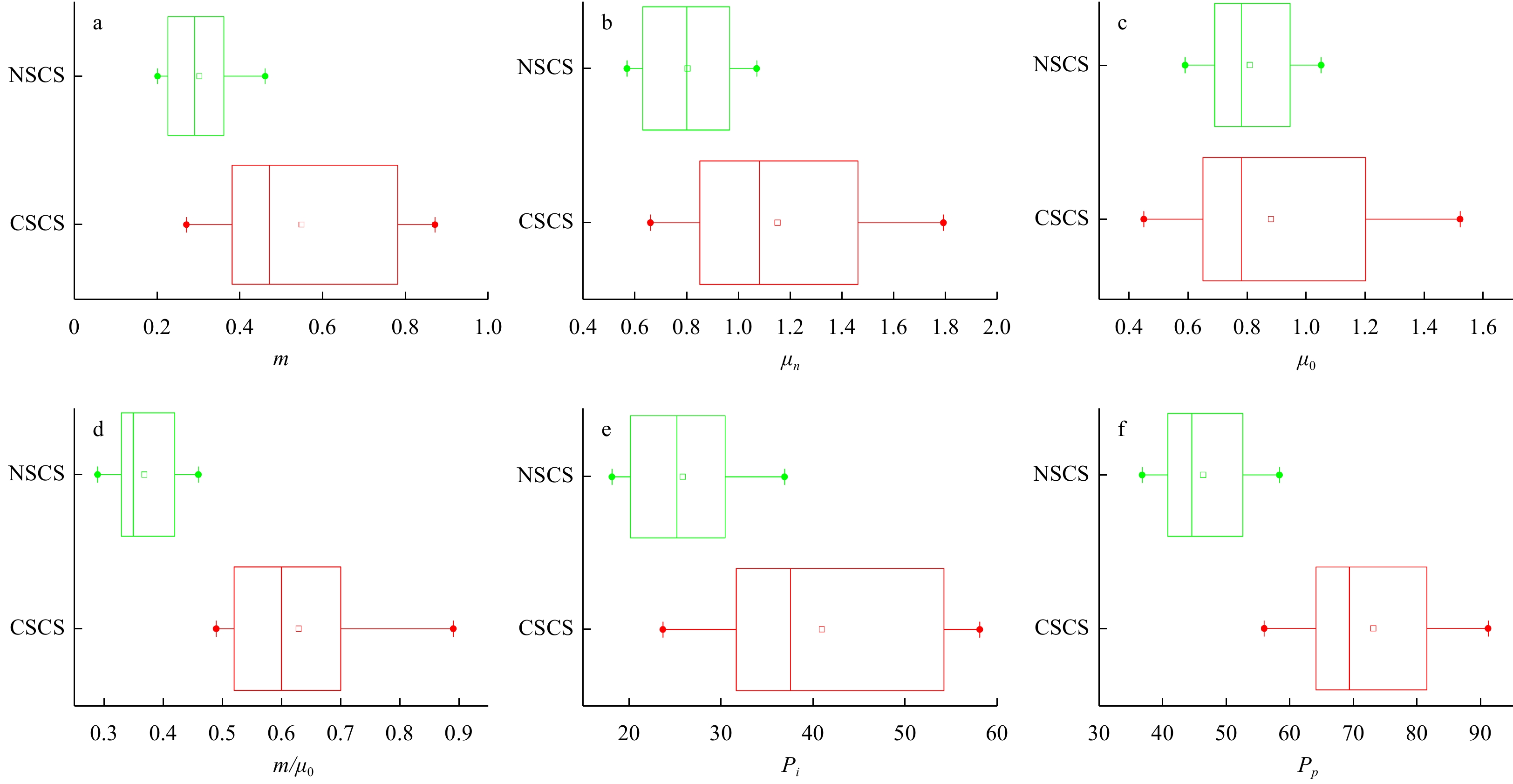
| Citation: | Dawei Chen, Congcong Guo, Linhui Yu, Yuanzhen Lu, Jun Sun. Phytoplankton growth and microzooplankton grazing in the central and northern South China Sea in the spring intermonsoon season of 2017[J]. Acta Oceanologica Sinica, 2020, 39(6): 84-95. doi: 10.1007/s13131-020-1593-1

|

| [1] |
Archer S D, Leakey R J G, Burkill P H, et al. 1996. Microbial dynamics in coastal waters of East Antarctica: herbivory by heterotrophic dinoflagellates. Marine Ecology Progress Series, 139: 239–255. doi: 10.3354/meps139239
|
| [2] |
Banse K. 2007. Do we live in a largely top-down regulated world?. Journal of Biosciences, 32(4): 791–796. doi: 10.1007/s12038-007-0080-6
|
| [3] |
Belzile C, Demers S, Ferreyra G A, et al. 2006. UV effects on marine planktonic food webs: a synthesis of results from mesocosm studies. Photochemistry and Photobiology, 82(4): 850–856. doi: 10.1562/2005-09-27-RA-699
|
| [4] |
Burkill P H, Mantoura R F C, Llewellyn C A, et al. 1987. Microzooplankton grazing and selectivity of phytoplankton in coastal waters. Marine Biology, 93(4): 581–590. doi: 10.1007/BF00392796
|
| [5] |
Calbet A, Landry M R. 2004. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnology and Oceanography, 49(1): 51–57. doi: 10.4319/lo.2004.49.1.0051
|
| [6] |
Caron D A, Dennett M R. 1999. Phytoplankton growth and mortality during the 1995 Northeast Monsoon and Spring Intermonsoon in the Arabian Sea. Deep Sea Research Part II: Topical Studies in Oceanography, 46(8–9): 1665–1690. doi: 10.1016/S0967-0645(99)00039-9
|
| [7] |
Caron D A, Hutchins D A. 2013. The effects of changing climate on microzooplankton grazing and community structure: drivers, predictions and knowledge gaps. Journal of Plankton Research, 35(2): 235–252. doi: 10.1093/plankt/fbs091
|
| [8] |
Chang J, Chung C C, Gong G C. 1996. Influences of cyclones on chlorophyll a concentration and Synechococcus abundance in a subtropical western Pacific coastal ecosystem. Marine Ecology Progress Series, 140: 199–205. doi: 10.3354/meps140199
|
| [9] |
Chen Bingzhang, Zheng Liping, Huang Bangqin, et al. 2013. Seasonal and spatial comparisons of phytoplankton growth and mortality rates due to microzooplankton grazing in the northern South China Sea. Biogeosciences, 10(4): 2775–2785. doi: 10.5194/bg-10-2775-2013
|
| [10] |
Chen Bingzhang, Landry M R, Huang Bangqin, et al. 2012. Does warming enhance the effect of microzooplankton grazing on marine phytoplankton in the ocean?. Limnology and Oceanography, 57(2): 519–526. doi: 10.4319/lo.2012.57.2.0519
|
| [11] |
Chen Bingzhang, Liu Hongbin, Landry M R, et al. 2009. Close coupling between phytoplankton growth and microzooplankton grazing in the western South China Sea. Limnology and Oceanography, 54(4): 1084–1097. doi: 10.4319/lo.2009.54.4.1084
|
| [12] |
Chen Y F L. 2005. Spatial and seasonal variations of nitrate-based new production and primary production in the South China Sea. Deep Sea Research Part I: Oceanographic Research Papers, 52(2): 319–340. doi: 10.1016/j.dsr.2004.11.001
|
| [13] |
Edwards E S, Burkill P H, Stelfox C E. 1999. Zooplankton herbivory in the Arabian Sea during and after the SW monsoon, 1994. Deep Sea Research Part II: Topical Studies in Oceanography, 46(3–4): 843–863. doi: 10.1016/S0967-0645(98)00130-1
|
| [14] |
First M R, Lavrentyev P J, Jochem F J. 2007. Patterns of microzooplankton growth in dilution experiments across a trophic gradient: implications for herbivory studies. Marine Biology, 151(5): 1929–1940. doi: 10.1007/s00227-007-0629-9
|
| [15] |
Gifford D J. 1988. Impact of grazing by microzooplankton in the Northwest Arm of Halifax Harbour, Nova Scotia. Marine Ecology Progress Series, 47: 249–258. doi: 10.3354/meps047249
|
| [16] |
Gifford D J, Fessenden L M, Garrahan P R, et al. 1995. Grazing by microzooplankton and mesozooplankton in the high-latitude North Atlantic Ocean: spring versus summer dynamics. Journal of Geophysical Research, 100(C4): 6665–6675. doi: 10.1029/94JC00983
|
| [17] |
Gómez F. 2007. Trends on the distribution of ciliates in the open Pacific Ocean. Acta Oecologica, 32(2): 188–202. doi: 10.1016/j.actao.2007.04.002
|
| [18] |
Gong G C, Liu K K, Liu C T, et al. 1992. The chemical hydrography of the South China Sea west of Luzon and a comparison with the West Philippine Sea. Terrestrial, Atmospheric and Oceanic Sciences, 3(4): 587–602. doi: 10.3319/TAO.1992.3.4.587(O)
|
| [19] |
Hansen H P, Koroleff F. 2007. Determination of nutrients. In: Grasshoff K, Kremling K, Ehrhardt M, eds. Methods of Seawater Analysis. 3rd ed. New York: Wiley
|
| [20] |
Hu Zifeng, Tan Yehui, Song Xingyu, et al. 2014. Influence of mesoscale eddies on primary production in the South China Sea during spring inter-monsoon period. Acta Oceanologica Sinica, 33(3): 118–128. doi: 10.1007/s13131-014-0431-8
|
| [21] |
Irigoien X, Flynn K J, Harris R P. 2005. Phytoplankton blooms: a ‘loophole’ in microzooplankton grazing impact?. Journal of Plankton Research, 27(4): 313–321. doi: 10.1093/plankt/fbi011
|
| [22] |
Jin Dexiang, Chen Jinhuan, Huang Kaige. 1965. China Marine Planktonic Diatoms (in Chinese). Shanghai: Shanghai Scientific and Technical Publishers, 1–219
|
| [23] |
Jyothibabu R, Devi C R A, Madhu N V, et al. 2008. The response of microzooplankton (20–200 μm) to coastal upwelling and summer stratification in the southeastern Arabian Sea. Continental Shelf Research, 28(4–5): 653–671. doi: 10.1016/j.csr.2007.12.001
|
| [24] |
Landry M R. 1993. Estimating rates of growth and grazing mortality of phytoplankton by the dilution method. In: Kemp P J, Cole J J, Sherr B F, et al., eds. Handbook of Methods in Aquatic Microbial Ecology. New York: CRC Press, 715–722
|
| [25] |
Landry M R, Brown S L, Campbell L, et al. 1998. Spatial patterns in phytoplankton growth and microzooplankton grazing in the Arabian Sea during monsoon forcing. Deep Sea Research Part II: Topical Studies in Oceanography, 45(10–11): 2353–2368. doi: 10.1016/S0967-0645(98)00074-5
|
| [26] |
Landry M R, Calbet A. 2004. Microzooplankton production in the oceans. ICES Journal of Marine Science, 61(4): 501–507. doi: 10.1016/j.icesjms.2004.03.011
|
| [27] |
Landry M R, Hassett R P. 1982. Estimating the grazing impact of marine micro-zooplankton. Marine Biology, 67(3): 283–288. doi: 10.1007/BF00397668
|
| [28] |
Lehman J T. 1991. Interacting growth and loss rates: the balance of top-down and bottom-up controls in plankton communities. Limnology and Oceanography, 36(8): 1546–1554. doi: 10.4319/lo.1991.36.8.1546
|
| [29] |
Leising A W, Horner R, Pierson J J, et al. 2005. The balance between microzooplankton grazing and phytoplankton growth in a highly productive estuarine fjord. Progress in Oceanography, 67(3–4): 366–383. doi: 10.1016/j.pocean.2005.09.007
|
| [30] |
Li Fengqi, Su Yusong. 2000. Analyses of Water Masses in Oceans (in Chinese). Qingdao: Qingdao Ocean University Press, 377–378
|
| [31] |
Liu Hongbin, Chang J, Tseng C M, et al. 2007. Seasonal variability of picoplankton in the Northern South China Sea at the SEATS station. Deep Sea Research Part II: Topical Studies in Oceanography, 54(14–15): 1602–1616. doi: 10.1016/j.dsr2.2007.05.004
|
| [32] |
Liu Hongbin, Suzuki K, Nishioka J, et al. 2009. Phytoplankton growth and microzooplankton grazing in the Sea of Okhotsk during late summer of 2006. Deep Sea Research Part I: Oceanographic Research Papers, 56(4): 561–570. doi: 10.1016/j.dsr.2008.12.003
|
| [33] |
Lonsdale D J, Caron D A, Dennett M R, et al. 2000. Predation by Oithona spp. on protozooplankton in the Ross Sea, Antarctica. Deep Sea Research Part II: Topical Studies in Oceanography, 47(15–16): 3273–3283. doi: 10.1016/S0967-0645(00)00068-0
|
| [34] |
López-Urrutia Á, Martin E S, Harris R P, et al. 2006. Scaling the metabolic balance of the oceans. Proceedings of the National Academy of Sciences of the United States of America, 103(23): 8739–8744. doi: 10.1073/pnas.0601137103
|
| [35] |
López-Urrutia Á. 2008. The metabolic theory of ecology and algal bloom formation. Limnology and Oceanography, 53(5): 2046–2047. doi: 10.4319/lo.2008.53.5.2046
|
| [36] |
Landry M R, Barber R T, Bidigare R R, et al. 1997. Iron and grazing constraints on primary production in the central equatorial Pacific: an EqPac Synthesis. Limnology and Oceanography, 42(3): 405–418. doi: 10.4319/lo.1997.42.3.0405
|
| [37] |
Macaluso A L, Mitchell D L, Sanders R W. 2009. Direct effects of UV-B radiation on the freshwater heterotrophic nanoflagellate Paraphysomonas sp. Applied and Environmental Microbiology, 75(13): 4525–4530. doi: 10.1128/AEM.02803-08
|
| [38] |
Nielsen T G, Kicrboe T. 1994. Regulation of zooplankton biomass and production in a temperate, coastal ecosystem. 2. Ciliates. Limnology and Oceanography, 39(3): 508–519. doi: 10.4319/lo.1994.39.3.0508
|
| [39] |
Ning X, Chai F, Xue H, et al. 2004. Physical-biological oceanographic coupling influencing phytoplankton and primary production in the South China Sea. Journal of Geophysical Research, 109(C10): C10005. doi: 10.1029/2004JC002365
|
| [40] |
Paranjape M A. 1987. Grazing by microzooplankton in the eastern Canadian arctic in summer 1983. Marine Ecology Progress Series, 40: 239–246. doi: 10.3354/meps040239
|
| [41] |
Porter K G, Sherr E B, Sherr B F, et al. 1985. Protozoa in planktonic food webs. The Journal of Protozoology, 32(3): 409–415. doi: 10.1111/j.1550-7408.1985.tb04036.x
|
| [42] |
Quevedo M, Viesca L, Anadón R, et al. 2003. The protistan microzooplankton community in the oligotrophicnorth-eastern Atlantic: large- and mesoscale patterns. Journal of Plankton Research, 25(5): 551–563. doi: 10.1093/plankt/25.5.551
|
| [43] |
Raven J A. 1998. The twelfth Tansley Lecture. Small is beautiful: the picophytoplankton. Functional Ecology, 12(4): 503–513
|
| [44] |
Rose J M, Caron D A. 2007. Does low temperature constrain the growth rates of heterotrophic protists? Evidence and implications for algal blooms in cold waters. Limnology and Oceanography, 52(2): 886–895. doi: 10.4319/lo.2007.52.2.0886
|
| [45] |
Schmoker C, Hernández-León S, Calbet A. 2013. Microzooplankton grazing in the oceans: impacts, data variability, knowledge gaps and future directions. Journal of Plankton Research, 35(4): 691–706. doi: 10.1093/plankt/fbt023
|
| [46] |
Setälä O, Kivi K. 2003. Planktonic ciliates in the Baltic Sea in summer: distribution, species association and estimated grazing impact. Aquatic Microbial Ecology, 32(3): 287–297
|
| [47] |
Shi Ping, Du Yan, Wang Dongxiao, et al. 2001. Annual cycle of mixed layer in South China Sea. Journal of Tropical Oceanography (in Chinese), 20(1): 10–17
|
| [48] |
Stelfox-Widdicombe C E, Edwards E S, Burkill P H, et al. 2000. Microzooplankton grazing activity in the temperate and sub-tropical NE Atlantic: summer 1996. Marine Ecology Progress Series, 208: 1–12. doi: 10.3354/meps208001
|
| [49] |
Strom S. 2002. Novel interactions between phytoplankton and microzooplankton: their influence on the coupling between growth and grazing rates in the sea. Hydrobiologia, 480(1–3): 41–54
|
| [50] |
Strom S L, Macri E L, Olson M B. 2007. Microzooplankton grazing in the coastal gulf of Alaska: variations in top-down control of phytoplankton. Limnology and Oceanography, 52(4): 1480–1494. doi: 10.4319/lo.2007.52.4.1480
|
| [51] |
Strom S L, Welschmeyer N A. 1991. Pigment-specific rates of phytoplankton growth and microzooplankton grazing in the open subarctic Pacific Ocean. Limnology and Oceanography, 36(1): 50–63. doi: 10.4319/lo.1991.36.1.0050
|
| [52] |
Su Jilan. 2004. Overview of the South China Sea circulation and its influence on the coastal physical oceanography outside the Pearl River Estuary. Continental Shelf Research, 24(16): 1745–1760. doi: 10.1016/j.csr.2004.06.005
|
| [53] |
Su Qiang, Huang Liangmin, Tan Yehui, et al. 2007. Preliminary study of microzooplankton grazing and community composition in the north of South China Sea in autumn. Marine Science Bulletin, 9(2): 43–53
|
| [54] |
Sun Jun. 2013. Carbon calculation on marine phytoplankton and its related fishery carbon sink. Progress in Fishery Sciences (in Chinese), 34(1): 90–96
|
| [55] |
Sun Jun, Feng Yuanyuan, Wang Dan, et al. 2013. Bottom-up control of phytoplankton growth in spring blooms in Central Yellow Sea, China. Deep Sea Research Part II: Topical Studies in Oceanography, 97: 61–71. doi: 10.1016/j.dsr2.2013.05.006
|
| [56] |
Sun Jun, Feng Yuanyuan, Zhang Yaohong, et al. 2007. Fast microzooplankton grazing on fast-growing, low-biomass phytoplankton: a case study in spring in Chesapeake Bay, Delaware Inland Bays and Delaware Bay. Hydrobiologia, 589(1): 127–139. doi: 10.1007/s10750-007-0730-6
|
| [57] |
Sun Jun, Guo Shujin. 2011. Dinoflagellate heterotrophy. Acta Ecologica Sinica (in Chinese), 31(20): 6270–6286
|
| [58] |
Tan Saichun, Shi Guangyu. 2009. Spatiotemporal variability of satellite-derived primary production in the South China Sea, 1998–2006. Journal of Geophysical Research, 114(G3): G03015
|
| [59] |
Teixeira I G, Figueiras F G. 2009. Feeding behaviour and non-linear responses in dilution experiments in a coastal upwelling system. Aquatic Microbial Ecology, 55(1): 53–63
|
| [60] |
Tian Yongqing, Huang Honghui, Gong Xiuyu, et al. 2016. The formation mechanism of the low temperature and high salinity water mass near the Zhongsha Islands in the South China Sea in March 2014. Journal of Tropical Oceanography (in Chinese), 35(2): 1–9
|
| [61] |
Tsai A Y, Chiang K P, Chang J, et al. 2005. Seasonal diel variations of picoplankton and nanoplankton in a subtropical western Pacific coastal ecosystem. Limnology and Oceanography, 50(4): 1221–1231. doi: 10.4319/lo.2005.50.4.1221
|
| [62] |
Utermöhl H. 1958. Zur vervollkommnung der quantitativen phytoplankton-methodik: mit 1 tabelle und 15 abbildungen im text und auf 1 tafel. SIL Communications, 9(1): 1–38
|
| [63] |
Verity P G, Stoecker D K, Sieracki M E, et al. 1993. Grazing, growth and mortality of microzooplankton during the 1989 North Atlantic spring bloom at 47°N, 18°W. Deep Sea Research Part I: Oceanographic Research Papers, 40(9): 1793–1814. doi: 10.1016/0967-0637(93)90033-Y
|
| [64] |
Verity P G, Wassmann P, Frischer M E, et al. 2002. Grazing of phytoplankton by microzooplankton in the Barents Sea during early summer. Journal of Marine Systems, 38(1–2): 109–123. doi: 10.1016/S0924-7963(02)00172-0
|
| [65] |
Welschmeyer N A. 1994. Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and pheopigments. Limnology and Oceanography, 39(8): 1985–1992. doi: 10.4319/lo.1994.39.8.1985
|
| [66] |
Wong G T F, Ku T L, Mulholland M, et al. 2007. The Southeast Asian Time-series Study (SEATS) and the biogeochemistry of the South China Sea—an overview. Deep Sea Research Part II: Topical Studies in Oceanography, 54(14–15): 1434–1447. doi: 10.1016/j.dsr2.2007.05.012
|
| [67] |
Wu Jingfeng, Chung S W, Wen L S, et al. 2003. Dissolved inorganic phosphorus, dissolved iron, and Trichodesmium in the oligotrophic South China Sea. Global Biogeochemical Cycles, 17(1): 1008
|
| [68] |
Yamaji I. 1966. Illustrations of the Marine Plankton of Japan. Osaka: Hoikusha Publishing Co Ltd, 1–538
|
| [69] |
Yang E J, Choi J K, Hyun J H. 2004. Distribution and structure of heterotrophic protist communities in the northeast equatorial Pacific Ocean. Marine Biology, 146(1): 1–15. doi: 10.1007/s00227-004-1412-9
|
| [70] |
Zhang Wuchang, Feng Meiping, Yu Ying, et al. 2012. Illustrated Guide to Contemporary Tintinnids in the World (in Chinese). Beijing: Science Press
|
| [71] |
Zhang Jiangtao, Yin Kedong, Dong Lihua. 2011. Microzooplankton grazing rate of size-fractionated phytoplankton in spring in the Yellow Sea, China. Marine Sciences (in Chinese), 35(9): 1–7
|
| [72] |
Zhang Wuchang, Yu Ying, Xiao Tian. 2015. An Illustrated Guide to Marine Planktonic Aloricate Oligotrich Ciliates (in Chinese). Beijing: Science Press
|
| [73] |
Zheng Guangming, Tang Danling. 2007. Offshore and nearshore chlorophyll increases induced by typhoon winds and subsequent terrestrial rainwater runoff. Marine Ecology Progress Series, 333: 61–74
|
| [74] |
Zhou Linbin, Tan Yehui, Huang Liangmin, et al. 2011. Phytoplankton growth and microzooplankton grazing in the continental shelf area of northeastern South China Sea after Typhoon Fengshen. Continental Shelf Research, 31(16): 1663–1671. doi: 10.1016/j.csr.2011.06.017
|
| [75] |
Zhou Linbin, Tan Yehui, Huang Liangmin, et al. 2015a. Does microzooplankton grazing contribute to the pico-phytoplankton dominance in subtropical and tropical oligotrophic waters?. Acta Ecologica Sinica, 35(1): 29–38. doi: 10.1016/j.chnaes.2014.12.007
|
| [76] |
Zhou Linbin, Tan Yehui, Huang Lianmin, et al. 2015b. Seasonal and size-dependent variations in the phytoplankton growth and microzooplankton grazing in the southern South China Sea under the influence of the East Asian monsoon. Biogeosciences, 12(22): 6809–6822. doi: 10.5194/bg-12-6809-2015
|
| [77] |
Zubkov M V, Sleigh M A, Burkill P H. 2000. Assaying picoplankton distribution by flow cytometry of underway samples collected along a meridional transect across the Atlantic Ocean. Aquatic Microbial Ecology, 21(1): 13–20
|

Figures(6) / Tables(7)

Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: