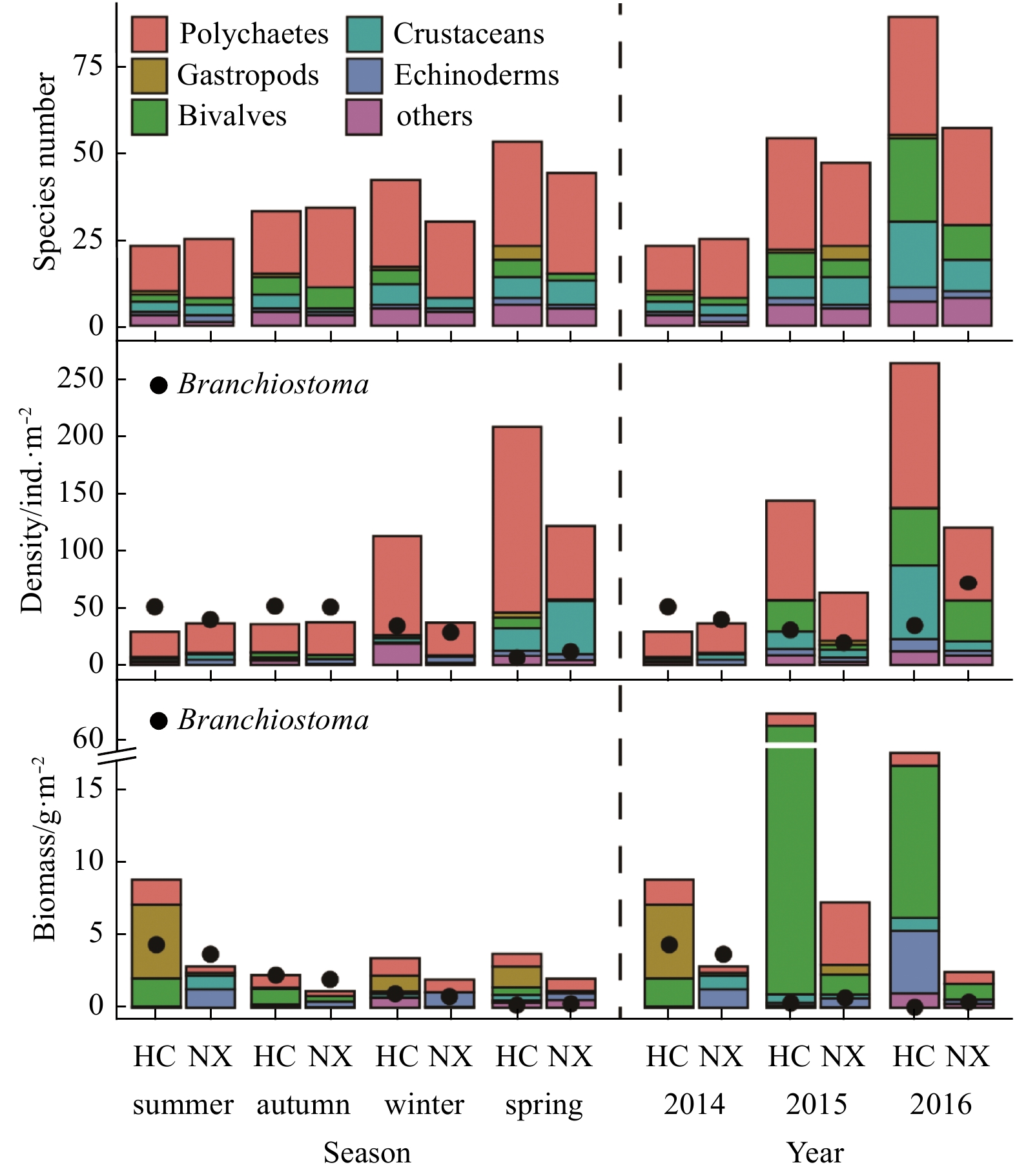
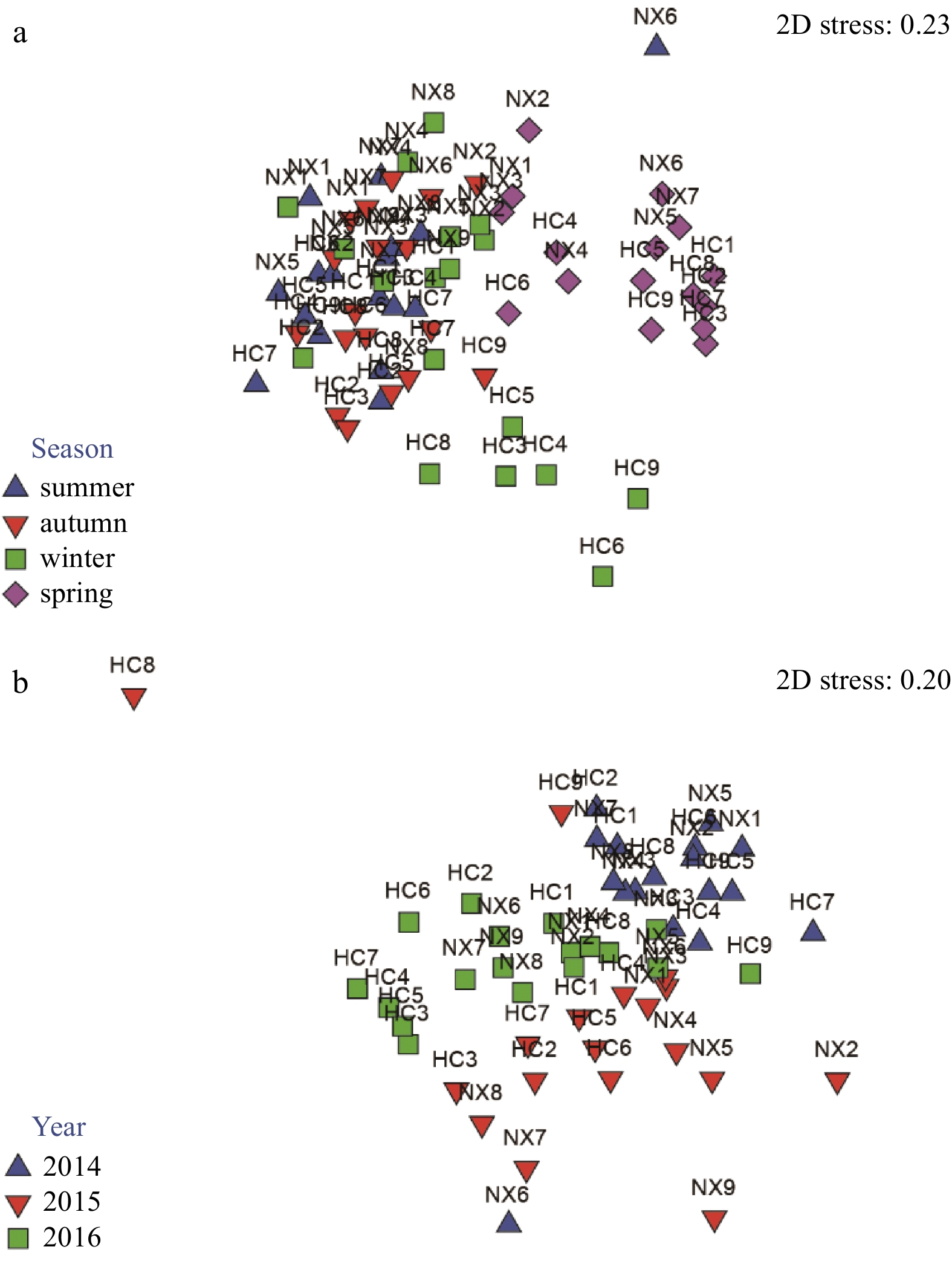
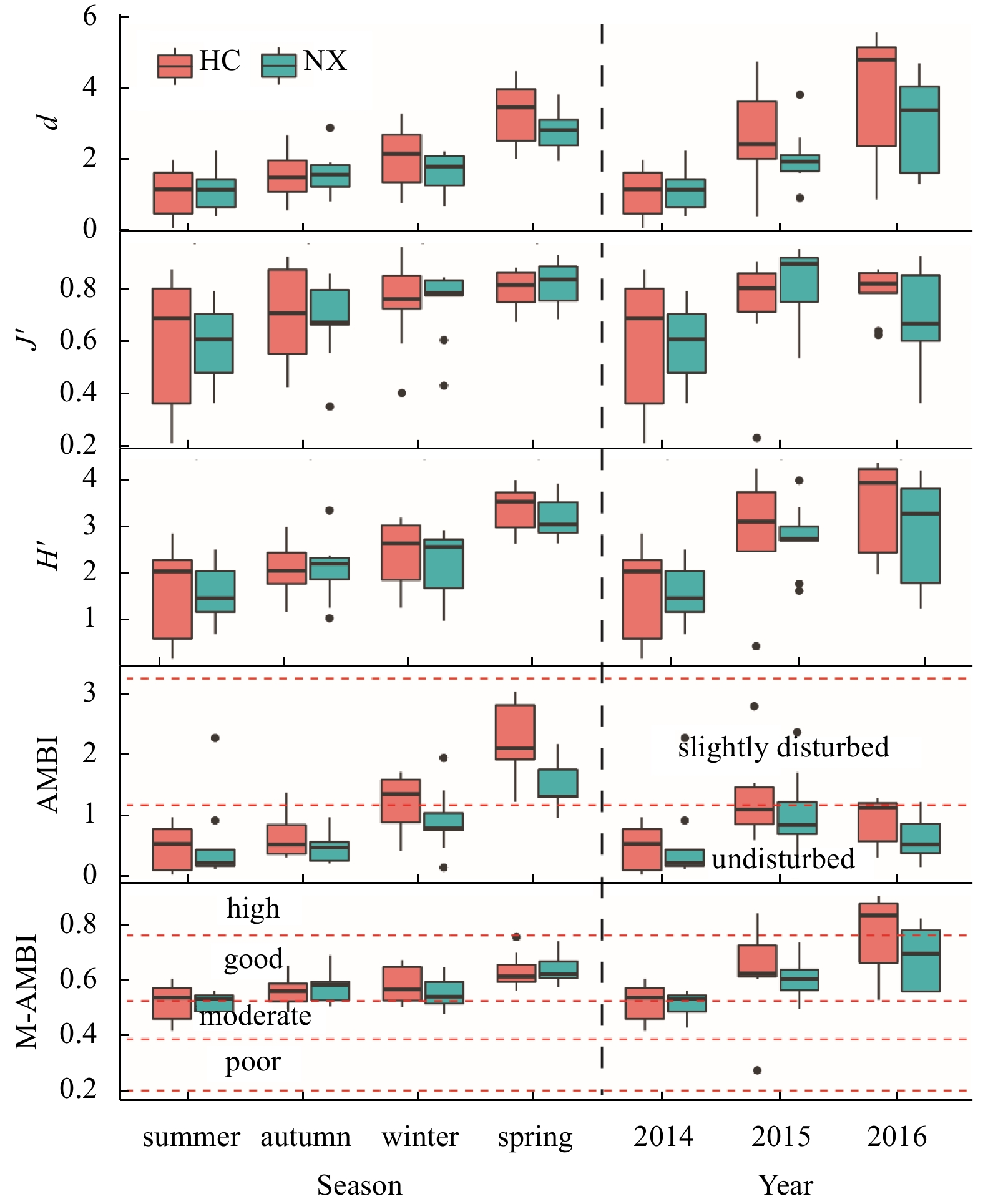
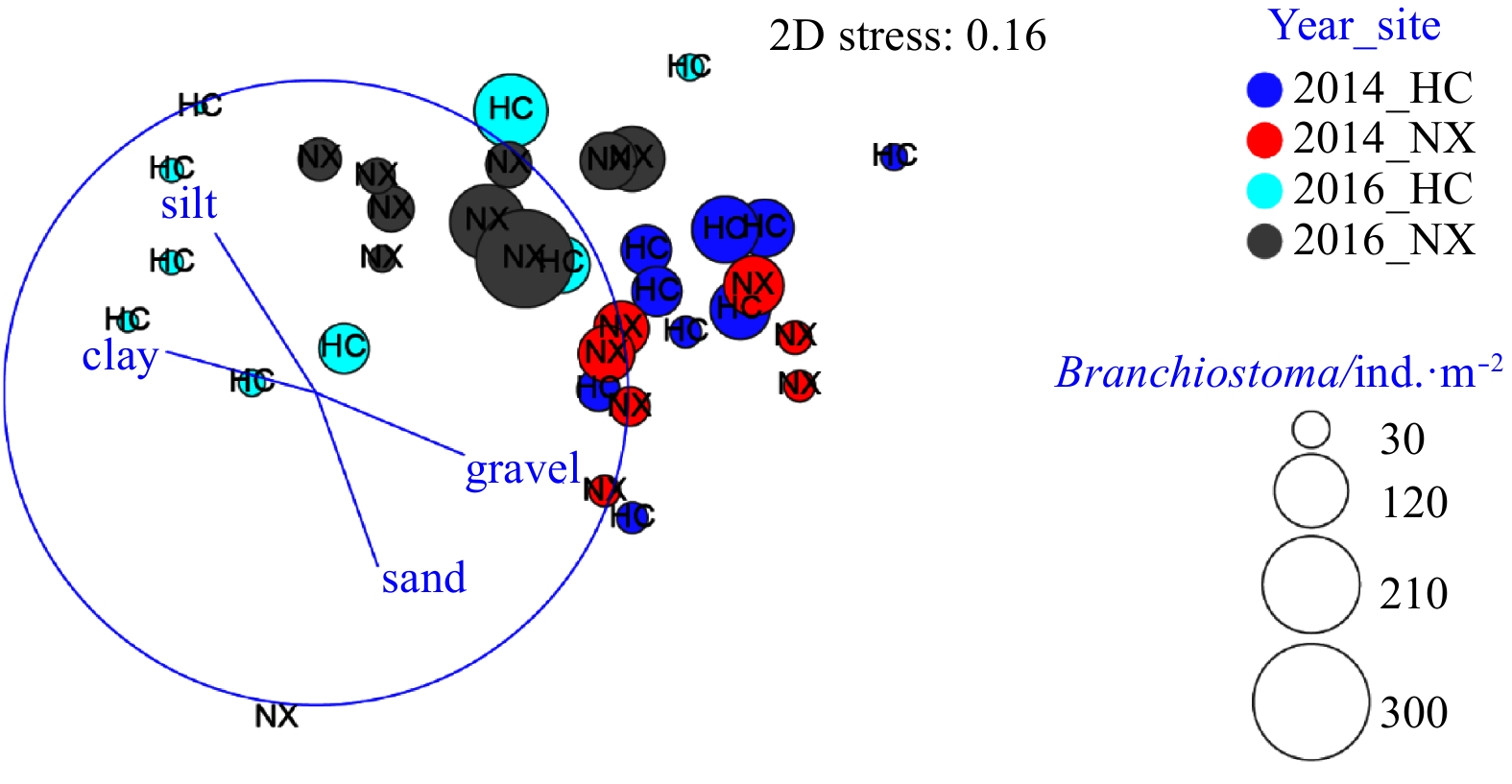
| Citation: | Yiyong Rao, Lizhe Cai, Wenjun Li, Xinwei Chen, Deyuan Yang. Spatiotemporal variations of benthic macrofaunal community in the Xiamen Amphioxus Nature Reserve, eastern South China Sea[J]. Acta Oceanologica Sinica, 2020, 39(6): 10-18. doi: 10.1007/s13131-020-1587-z

|

| [1] |
Antoniadou C, Krestenitis Y, Chintiroglou C. 2004. Structure of the “Amphioxus Sand” community in Thermaikos Bay (Eastern Mediterranean). Fresenius Environmental Bulletin, 13(11): 1122–1128
|
| [2] |
Barnes R S K. 2016. Spatial homogeneity of benthic macrofaunal biodiversity across small spatial scales. Marine Environmental Research, 122: 148–157. doi: 10.1016/j.marenvres.2016.10.006
|
| [3] |
Bertrán C, Fierro P, Encalada E, et al. 2016. Macrobenthos of the coastal Budi Lagoon, southern Chile: Changes associated with seasonal environmental variation. Brazilian Journal of Oceanography, 64(3): 239–248. doi: 10.1590/S1679-87592016113306403
|
| [4] |
Bessa F, Gonçalves S C, Franco J N, et al. 2014. Temporal changes in macrofauna as response indicator to potential human pressures on sandy beaches. Ecological Indicators, 41(2): 49–57
|
| [5] |
Borja A, Dauer D M. 2008. Assessing the environmental quality status in estuarine and coastal systems: Comparing methodologies and indices. Ecological Indicators, 8(4): 331–337. doi: 10.1016/j.ecolind.2007.05.004
|
| [6] |
Borja A, Franco J, Pérez V. 2000. A marine Biotic Index to establish the ecological quality of soft-bottom benthos within European estuarine and coastal environments. Marine Pollution Bulletin, 40(12): 1100–1114. doi: 10.1016/S0025-326X(00)00061-8
|
| [7] |
Cai Lizhe, Lin Junda, Li Hongmei. 2001. Macroinfauna communities in an organic-rich mudflat at Shenzhen and Hong Kong, China. Bulletin of Marine Science (in Chinese), 69(3): 1129–1138
|
| [8] |
Chen Y, Cheung S G, Shin P K S. 2013. A baseline study of benthic community associated with Amphioxus Sand in subtropical Hong Kong. Marine Pollution Bulletin, 72(1): 274–280. doi: 10.1016/j.marpolbul.2013.03.026
|
| [9] |
Clarke K R, Gorley R N, Somerfield P, et al. 2014. Change in marine communities: an approach to statistical analysis and interpretation. 3rd ed. Plymouth: Primer-E Ltd
|
| [10] |
Dauer D M, Ranasinghe J A, Weisberg S B. 2000. Relationships between benthic community condition, water quality, sediment quality, nutrient loads, and land use patterns in Chesapeake Bay. Estuaries, 23(1): 80–96. doi: 10.2307/1353227
|
| [11] |
De Moura Barboza C A, Hadlich H L, Sandrini-Neto L, et al. 2013. Is the distribution of the lancelet Branchiostoma caribaeum affected by sewage discharges? An analysis at multiple scales of variability. Marine Pollution Bulletin, 69(1–2): 178–188. doi: 10.1016/j.marpolbul.2013.01.040
|
| [12] |
Dean H K. 2008. The use of polychaetes (annelida) as indicator species of marine pollution: a review. Revista De Biología Tropical, 56(4): 11–38
|
| [13] |
Díaz-Tapia P, Bárbara I, Díez I. 2013. Multi-scale spatial variability in intertidal benthic assemblages: differences between sand-free and sand-covered rocky habitats. Estuarine Coastal and Shelf Science, 133: 97–108. doi: 10.1016/j.ecss.2013.08.019
|
| [14] |
Dubois S, Gelpi Jr C G, Condrey R E, et al. 2009. Diversity and composition of macrobenthic community associated with sandy shoals of the Louisiana continental shelf. Biodiversity and Conservation, 18(14): 3759–3784. doi: 10.1007/s10531-009-9678-3
|
| [15] |
Dutertre M, Hamon D, Chevalier C, et al. 2013. The use of the relationships between environmental factors and benthic macrofaunal distribution in the establishment of a baseline for coastal management. ICES Journal of Marine Science, 70(2): 294–308. doi: 10.1093/icesjms/fss170
|
| [16] |
Fang Shaohua, Lü Xiaomei, Zhang Yueping. 2002. Resources and protection of amphioxus (Branchiostoma belcheri) in Xiamen National Natural Reserve Area of Rare Marine Species. Marine Sciences (in Chinese), 26(10): 9–12
|
| [17] |
França S, Vinagre C, Pardal M A, et al. 2009. Spatial and temporal patterns of benthic invertebrates in the Tagus estuary, Portugal: comparison between subtidal and an intertidal mudflat. Scientia Marina, 73(2): 307–318. doi: 10.3989/scimar.2009.73n2307
|
| [18] |
Glockzin M, Zettler M L. 2008. Spatial macrozoobenthic distribution patterns in relation to major environmental factors—A case study from the Pomeranian bay (southern Baltic Sea). Journal of Sea Research, 59(3): 144–161. doi: 10.1016/j.seares.2008.01.002
|
| [19] |
Higgins R P, Kristensen R M. 1986. New Loricifera from southeastern United States coastal waters. Smithsonian Contributions to Zoology, 438: 1–70
|
| [20] |
Hu Min. 2013. The reproductive cycle of Branchiostoma belcheri and B. japonicum in Xiamen Waters (in Chinese) [dissertation]. Fuzhou: Fujian Normal University
|
| [21] |
Huang Liangmin, Li Zhongbao, Lin Tao, et al. 2013. Assessment on Branchiostoma belcheri stock health in Xiamen National Rare Marine Species Nature Reserve. Oceanologia et Limnologia Sinica (in Chinese), 44(1): 103–110
|
| [22] |
Jan S, Wang J, Chern C S, et al. 2002. Seasonal variation of the circulation in the Taiwan Strait. Journal of Marine Systems, 35(3–4): 249–268. doi: 10.1016/S0924-7963(02)00130-6
|
| [23] |
Johansen P O, Isaksen T E, Bye-Ingebrigtsen E, et al. 2018. Temporal changes in benthic macrofauna on the west coast of Norway resulting from human activities. Marine Pollution Bulletin, 128: 483–495. doi: 10.1016/j.marpolbul.2018.01.063
|
| [24] |
Karalis P, Antoniadou C, Chintiroglou C. 2003. Structure of the artificial hard substrate assemblages in ports in Thermaikos Gulf (North Aegean Sea). Oceanologica Acta, 26(3): 215–224. doi: 10.1016/S0399-1784(03)00040-9
|
| [25] |
Konsulova T. 1992. Seasonal structure and ecological status of Varna Bay (Black Sea) sandy and muddy macrozoobenthic coenoses. Rapp Com Mer Meditt, 33: 42
|
| [26] |
Lancellotti D A, Stotz W B. 2004. Effects of shoreline discharge of iron mine tailings on a marine soft-bottom community in northern Chile. Marine Pollution Bulletin, 48(3–4): 303–312. doi: 10.1016/j.marpolbul.2003.08.005
|
| [27] |
Levin S A. 1992. The problem of pattern and scale in ecology: the Robert H. MacArthur award lecture. Ecology, 73(6): 1943–1967
|
| [28] |
Light S F. 1923. Amphioxus fisheries near the University of Amoy, China. Science, 58(1491): 57–60. doi: 10.1126/science.58.1491.57
|
| [29] |
Lin Hsiuchin, Chen Jenping, Chan B K K, et al. 2015. The interplay of sediment characteristics, depth, water temperature, and ocean currents shaping the biogeography of lancelets (Subphylum Cephalochordata) in the NW Pacific waters. Marine Ecology, 36(3): 780–793. doi: 10.1111/maec.12183
|
| [30] |
Ma Zhijun, Chen Ying, Melville D S, et al. 2019. Changes in area and number of nature reserves in China. Conservation Biology, 33(5): 1066–1075. doi: 10.1111/cobi.13285
|
| [31] |
Mann K H, Lazier J R N. 2006. Dynamics of Marine Ecosystems. Biological-physical Interactions in the Oceans. 3rd ed. Cambridge: Blackwell Publishing, 503
|
| [32] |
Olsson J, Bergström L, Gårdmark A. 2013. Top-down regulation, climate and multi-decadal changes in coastal zoobenthos communities in two Baltic Sea areas. PLoS One, 8(5): e64767. doi: 10.1371/journal.pone.0064767
|
| [33] |
Ramey P A, Snelgrove P V R. 2003. Spatial patterns in sedimentary macrofaunal communities on the south coast of Newfoundland in relation to surface oceanography and sediment characteristics. Marine Ecology Progress Series, 262(5): 215–227
|
| [34] |
Rao Yiyong, Zhang Huiping, Lin Dan, et al. 2015. Research on the status of Amphioxus and other macrofauna resources in Xiamen waters. Marine Science Bulletin (in Chinese), 34(5): 577–585
|
| [35] |
Rosenberg R, Magnusson M, Nilsson H C. 2009. Temporal and spatial changes in marine benthic habitats in relation to the EU water framework directive: the use of sediment profile imagery. Marine Pollution Bulletin, 58(4): 565–572. doi: 10.1016/j.marpolbul.2008.11.023
|
| [36] |
Rota E, Perra G, Focardi S. 2009. The European lancelet Branchiostoma lanceolatum (Pallas) as an indicator of environmental quality of Tuscan Archipelago (Western Mediterranean Sea). Chemistry and Ecology, 25(1): 61–69. doi: 10.1080/02757540802641361
|
| [37] |
Schückel U, Beck M, Kröncke I. 2015. Macrofauna communities of tidal channels in Jade Bay (German Wadden Sea): spatial patterns, relationships with environmental characteristics, and comparative aspects. Marine Biodiversity, 45(4): 841–855. doi: 10.1007/s12526-014-0308-2
|
| [38] |
Seitz R D, Knick K E, Davenport T M, et al. 2018. Human influence at the coast: upland and shoreline stressors affect coastal macrofauna and are mediated by salinity. Estuaries and Coasts, 41(1): 114–130. doi: 10.1007/s12237-017-0274-6
|
| [39] |
Shahidul I M, Tanaka M. 2004. Impacts of pollution on coastal and marine ecosystems including coastal and marine fisheries and approach for management: a review and synthesis. Marine Pollution Bulletin, 48(7–8): 624–649. doi: 10.1016/j.marpolbul.2003.12.004
|
| [40] |
Shin P K S, Huang Z G, Wu R S S. 2004. An updated baseline of subtropical macrobenthic communities in Hong Kong. Marine Pollution Bulletin, 49(1–2): 128–135
|
| [41] |
Silva L F B D, Tavares M, Soares-Gomes A. 2008. Population structure of the lancelet Branchiostoma caribaeum (Cephalochordata: Branchiostomidae) in the Baía de Guanabara, Rio de Janeiro, southeastern Brazil. Revista Brasileira De Zoologia, 25(4): 617–623. doi: 10.1590/S0101-81752008000400006
|
| [42] |
Spencer B E, Kaiser M J, Edwards D B. 1997. Ecological effects of intertidal Manila clam cultivation: observations at the end of the cultivation phase. Journal of Applied Ecology, 34(2): 444–452. doi: 10.2307/2404889
|
| [43] |
Thrush S F, Lundquist C J, Hewitt J E. 2005. Spatial and temporal scales of disturbance to the seafloor: a generalized framework for active habitat management. American Fisheries Society Symposium, 41: 639–649
|
| [44] |
Varfolomeeva M, Naumov A. 2013. Long-term temporal and spatial variation of macrobenthos in the intertidal soft-bottom flats of two small bights (Chupa Inlet, Kandalaksha Bay, White Sea). Hydrobiologia, 706: 175–189. doi: 10.1007/s10750-012-1341-4
|
| [45] |
Veiga P, Redondo W, Sousa-Pinto I, et al. 2017. Relationship between structure of macrobenthic assemblages and environmental variables in shallow sublittoral soft bottoms. Marine Environmental Research, 129: 396–407. doi: 10.1016/j.marenvres.2017.07.002
|
| [46] |
Veiga P, Torres A C, Aneiros F, et al. 2016. Consistent patterns of variation in macrobenthic assemblages and environmental variables over multiple spatial scales using taxonomic and functional approaches. Marine Environmental Research, 120: 191–201. doi: 10.1016/j.marenvres.2016.08.011
|
| [47] |
Vergara M, Oliva M E, Riascos J M. 2012. Population dynamics of the amphioxus Branchiostoma elongatum from northern Chile. Journal of the Marine Biological Association of the United Kingdom, 92(3): 591–599. doi: 10.1017/S0025315411000804
|
| [48] |
Wang Weiyang, Chen Bizhe, Yao Lianteng, et al. 1989. Investigation on the stock of lancelet (Branchiostoma belcheri Gray) in the shallow sea area at qianpu, Xiamen. Fujian Fisheries (in Chinese), (1): 17–22
|
| [49] |
Wang Lu, Fan Ying, Yan Cunjun, et al. 2017. Assessing benthic ecological impacts of bottom aquaculture using macrofaunal assemblages. Marine Pollution Bulletin, 114(1): 258–268. doi: 10.1016/j.marpolbul.2016.09.032
|
| [50] |
Weng Zhaohong, Xie Yangjie, Xiao Zhiqun, et al. 2012. The amphioxus (Branchiostoma belcheri (Gray)) resources status and its ecological and environmental assessment in Xiamen Coast. Journal of Jimei University (Natural Science) (in Chinese), 17(4): 259–264
|
| [51] |
Weng Zhaohong, Zhang Yazhi, Liu Xiande, et al. 2010. Lancelet distribution in Fujian sea waters and its protection strategy. Marine Sciences (in Chinese), 34(8): 35–40
|
| [52] |
Zeng Guoshou, He Minhai, Cheng Zhaodi. 1996. Monitoring and research in lancelet protected area in Huangcuo, Xiamen. Journal of Oceanography in Taiwan Strait (in Chinese), 15(2): 174–181
|
| [53] |
Zhang Qiujin, Zhong Jing, Fang Shaohua, et al. 2006. Branchiostoma japonicum and B. belcheri are distinct lancelets (Cephalochordata) in Xiamen waters in China. Zoological Science, 23(6): 573–579. doi: 10.2108/zsj.23.573
|
| [54] |
Zhou Qiulin, He Minghai, Shao Hedao, et al. 1990. Distribution of lancelet and its relationship to bottom soil in Huangcuo waters of Xiamen. Journal of Oceanography in Taiwan Strait (in Chinese), 9(2): 185–189
|
| [55] |
Zinn D J. 1968. A brief consideration of the current terminology and sampling procedures used by investigators of marine interstitial fauna. Transactions of the American Microscopical Society, 87(2): 219–225. doi: 10.2307/3224445
|
Figures(5) / Tables(3)

Supported by:
Beijing Renhe Information Technology Co. Ltd





 DownLoad:
DownLoad: