Key Laboratory of Marine Bioactive Substances, First Institute of Oceanography, Ministry of Natural Resources, Qingdao 266061, China
2.
Laboratory for Marine Biology and Biotechnology, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266237, China
Funds:
The Marine Public Welfare Project of SOA under contract No. 201505032; the Scientific and Technological Innovation Project financially of Qingdao National Laboratory for Marine Science and Technology under contract No. 2016ASKJ14.
Seven bacterial clones with alginate-utilizing activity were isolated from rotten kelp. By activity test, the Vibrio sp. QD-5 with the potential alginate-degrading capability was chosen to carry out the draft genome sequencing, and the result showed that the Vibrio sp. QD-5 containing an alginate lyase gene cluster. One of these genes, aly-IV, was cloned and characterized for the first time. After overexpression, Aly-IV, with a molecular mass of about 62 kDa and a theoretical isoelectric point (pI) of 5.12, was purified to a specific activity of 1 256.78 U/mg and showed highest activity at 35°C in the Tris-HCl buffer at pH of 8.9. Moreover, the enzyme activity was enhanced by the metal ions of Na+, K+ and Mg2+ under certain concentration. Aly-IV degraded favorably polyG blocks in an endo-type, yielding monomer and dimer as the main products. Due to its high substrate specificity, Aly-IV could be used as a potential tool for production of polyG oligosaccharides with low degree of polymerization (DP) and for determining the fine structure of alginate.
Alginates are quite abundant polysaccharide in nature as structural components in marine brown algae (Phaeophyceae) and as capsular polysaccharides in soil bacteria (Gacesa, 1992), which consists of β-D-mannuronate (M) and α-L-guluronate (G) as monomeric units. Commercial alginates are mostly produced by the brown algae such as Laminaria hyperborea,Macrocystis pyrifera, and Laminaria japonica. Pharmaceutical, food and technical applications (such as in print paste for the textile industry) are the quantitative main market fields for alginates. Alginate oligosaccharides have attracted increasing attention because of their wide applications in food and pharmaceutical industry (Fremond et al., 1993; Steinlechner-Maran et al., 1996; Atala et al., 1994). They can be used as growth promoters for plants and therapeutic agents such as anticoagulants and tumor inhibitors (Hu et al., 2004; Iwamoto et al., 2005; An et al., 2009). They can also induce the cytokine production and regulate the blood sugar as well as lipid (Zhang et al., 2004; Zong et al., 2014). With the high-efficiency and high-specificity, alginate lyases, especially endolytic alginate lyases, have been widely used in the production of alginate oligosaccharides (Li et al., 2011), the elucidation of the fine structures of alginate (Aarstad et al., 2012; Boyd and Turvey, 1978; Østgaard, 1993) and the preparation of protoplast of red and brown algae (Inoue et al., 2011; Hu et al., 2003; Butler et al., 1989). Alginate lyases have been prepared from various sources, including marine algae, marine mollusks, marine bacteria, soil bacteria, fungi and viruses (Wong et al., 2000). These lyases were used to break the glycoside bond in alginate through β–elimination mechanism to form a double band between C4 and C5 at the non–reducing end of the degrading product (Wong et al., 2000). According to their different substrate specificities, alginate lyases were categorized into M–specific lyase (EC 4.2.2.3), G–specific lyase (EC 4.2.2.11). But until now, most of the reported alginate lyases were M–specific lyase. Based on their primary structures, the alginate lyases were also grouped into seven polysaccharide lyase families (PLs) in the Carbohydrate Active Enzyme (CAZy) database (http://www.cazy.org/), including PL–5, PL–6, PL–7, PL–14, PL–15, PL–17 and PL–18 (Huang et al., 2013). Most alginate lyases displayed in the CAZy database were assigned to PL7 accounting for more than 40%.
In this study, an alginate lyase–producing bacteria Vibrio sp. QD-5 was isolated from rotten kelp. The alginate lyase gene (aly–IV) was cloned from this strain and expressed in Escherichia coli. The recombinant enzyme Aly–IV possess the polyG–specific alginate lyase activity, and its enzymatic properties were characterized.
2.
Materials and methods
2.1
Screening and identification of alginate-degrading bacteria
Alginate-degrading bacteria were isolated from the rotten kelp. The decaying kelp was taken out and placed in sterile water to homogenize. The homogenate was diluted with deionized water serially and then spread on agar plate containing 0.5% (w/v) sodium alginate, 0.5% (w/v) (NH4)2SO4, 0.2% (w/v) K2HPO4, 0.1% (w/v) MgSO4, 0.001% (w/v) FeSO4·7H2O, and 2% (w/v) agar power. The plates were incubated at 28°C for about 72 h and plenty of colonies grew on the plate. One colony named QD-5 with highest intracellular alginate lyase activity was selected and cultured in 2216E media with alginate or Luria–Bertani (LB) broth for the follow-up study. The 16S rDNA sequences of the strains were amplified from the genomic DNA by PCR. The 16S rRNA genes were blasted using nucleotide BLAST in the NCBI and aligned with its closely related sequences retrieved from GenBank database using DNAMAN. The phylogenetic tree was constructed through neighbor–joining algorithms.
2.2
Cloning and sequence analysis of the alginate lyase gene
In order to obtain an alginate lyase gene, the genome of QD-5 was sequenced. The genome annotation reveals that an open reading frame coding for complex polysaccharide depolymerases was predicted. Using the designed primers P1 (5′–TAGGTGGGATCCGATCATGAAGC–3′) and P2 (5′–AGAACTCGAGTTGGTACTTACC–3′), the gene named aly-IV was amplified, purified and sequenced. The ORFs (open reading frames) of the DNA sequence were translated into corresponding amino acid sequences using the translate tool on the ExPASy sever (http://web.expasy.org/translate/). Various physical and chemical parameters of the predicted protein were calculated using ProtParam (http://web.expasy.org/protparam/). Functional information and the similarity sequences of the putative protein were performed on NCBI (https://www.ncbi.nlm.nih.gov/). Protein domains were identified using the SMART (http://smart.embl-heidelberg.de/).
2.3
Production of recombinant Aly-IV
For express Aly-IV, the DNA fragment were digested by BamHI and XhoI, and then ligated into the BamHI and XhoI sites of pET–22b (+). The resultant plasmids were transferred into E. coil BL21 (DE3) cell. The E. coli BL21–pET–22b (+)–aly–IV was cultured in LB broth supplemented with 100 μg/mL ampicillin at 37°C until OD600 reached 0.6–0.8, the culture was supplemented with isopropyl-β-D-thiogalactopyranoside (IPTG) at a final concentration of 1 mmol/L to induce the expression of Aly–IV. The cells were cultured for another 6 h at 37°C, 200 r/min and harvested by centrifugation. To optimize expression, a series of parameters including different IPTG concentrations, post induction temperature and time were tested. However, the target protein still existed in the form of inclusion body (IB).
2.4
Purification of Aly-IV
As Tao et al. (2010) described before, the inclusion body was solubilized by sarkosyl. The prepared dissolved inclusion body solution was subjected to Ni-NTA Sepharose column (GE Healthcare, Uppsala, Sweden) pre-equilibrated with binding buffer (20 mmol/L Na3PO4·12H2O, 0.5 mol/L NaCl, 10 mmol/L imidazole, pH 7.4). The fractions containing most of the target enzyme were eluted using elution buffer (20 mmol/L Na3PO4·12H2O, 0.5 mol/L NaCl, 200 mmol/L imidazole, pH 7.4) and pooled for the further purified using Superdex-200 Increase10/300 (GE Healthcare, Uppsala, Sweden). The enzyme was eluted with Tris–HCl buffer (pH 8.6) at a flow rate of 0.15 mL/min. Fractions with alginate lyase activity were gathered and stored at –20°C. The purity of the fractions and the molecular weight of the enzyme were determined by SDS–PAGE. The concentration of the protein was measured by Coomassie Brilliant Blue G–250 with bovine serum albumin (BSA) as standard.
2.5
Assay of the recombinant alginate lyase Activity
The activity of the alginate lyase Aly–IV was measured in a mixture solution containing 0.3% (w/v) sodium alginate solubilized in 50 mmol/L Tris–HCl pH 8.9 (0.9 mL) and a suitably enzyme solution (0.1 mL) at 35°C for 10 min. The reaction was terminated in boiling water for 10 min. The enzyme activity was determined as the increase in the absorbance 235 nm. One unit (U) was defined as the amount of enzyme required to increase by 0.1 the absorbance at 235 nm per minute.
2.6
Enzymatic degradation of sodium alginate
A total of 0.1 mL 0.5% sodium alginate, 0.05 mL of enzyme and 0.15 mL 50 mmol/L Tris-HCl were mixed in 1.5 mL tube, and then the mixture was incubated at 35°C. After incubated for 0.5, 1, 2, 3, 4, 5, 6, 12, 24 and 36 h, the samples were taken to determine the amount of reducing sugars by using the 3, 5–dinitrosalicylic acid (DNS) colorimetry (Hou et al., 2009).
2.7
Substrate specificity and kinetic parameters of the recombinant enzyme
A total of 50 μL purified enzyme was added to the reaction mixture containing 0.5% of sodium alginate, polyG, polyM and polyGM, respectively, for the substrate specificity. Enzyme activity was measured as described previously.
Kinetic parameters of the enzyme towards sodium alginate, polyG, polyM and polyGM, were obtained by measuring the enzyme activity with substrate of different concentrations (0.1–1 g/mL) (Table 1). The initial velocities were measured in the first five minute in the standard assay mixture. The Km and Vmax values were analyzed using the Eadie–Hofstee method.
Table
1.
Specific activity and kinetic parameters of the recombinant enzyme Aly-IV towards sodium alginate, polyG, polyM and polyGM
Substrate
Km/mg∙mL–1
Vmax/U∙mg–1
Relative activity/%
Sodium alginate
0.22
1 056.8
100
polyG
0.33
1 247.6
68.41
polyM
2.89
506.3
0.53
polyGM
1.42
711.8
1.23
Note: The data were the means of three experimental repeats with SD<0.05. The initial reaction velocity was defined as the increasing absorbance at 235 nm of the products produced by recombinant enzyme Aly-IV at the first 5 min.
2.8
Characterization of the recombinant alginate lyase
The temperature effects on the enzyme towards sodium alginate were performed at pH 8.9 in different temperature of 10–55°C. The thermal stability of the enzyme towards sodium alginate was determined by measuring the enzyme activity after incubation at 10–55°C for 30 min. The pH effects on the enzyme towards sodium alginate were performed at 35°C at pHs ranging 5–10.5. The pH stability of the enzyme towards sodium alginate was determined by measuring the residual enzyme activity after incubated at different pH for 3 h in ice-bath.
The influences of NaCl on the enzyme were performed by adding different concentration of NaCl (1–45 mmol/L) to the reaction buffer. The enzyme activity without NaCl served as control. The effects of metal ions on the enzyme were carried out by measuring the enzyme activity in the presence of metal ions. The reaction mixture without any metal ion was taken as control.
2.9
Analysis of reaction mode and products
The degraded products of the recombinant enzyme Aly–IV were analyzed using thin–layer chromatography (TLC) (TLC silica gel 60 F254, Merck KGaA, Darmstadt, Germany). The digested samples were periodically withdrawn for TLC analysis. The products on the TLC plates were developed using a solvent system of 1–butanol/ethanol/water 2:1:1 and then visualized by heating the plates at 85°C for 10 min after spraying with a diphenylamine/aniline/phosphate reagent. To further determine the products of the enzymatic action, the ESI–TOF–MS was conducted. Five microliters samples with 6 h reaction time were loop-injected to a G1969A time of flight mass spectrometer (TOF–MS) (Agilent Technologies Inc., California, USA) after filtration with 0.22 μm filter membrane. The products were detected in a negative–ion mode using the following settings: pressure of the atomizer (N2), 40 psi; flow rate of the dryer (N2), 11 L/min; temperature of the dryer (N2), 350°C; capillary voltage, 4 000 V; scanning the mass range, 100–1 000 m/z.
To determine the reaction mode of the recombinant enzyme Aly–IV, the viscometric parameters were measured with a rotary viscosimeter (SNB-1; Sunny Hengping Scientific instrument Co., Ltd, Shanghai, China). The reaction mixtures contained 3 mL enzyme Aly–IV and 2 mL 1.5% sodium alginate dissolved in 50 mmol/L Tris–HCl, pH 8.9. The viscosity was measured at 35°C for timed interval and the timed samples (0.3 mL) were taken out for the measurement of the absorbance.
2.10
Genome sequence accession numbers
The genome sequence have been deposited in GenBank under the accession number PRJNA382465.
3.
Results and discussion
3.1
Screening of alginate-degrading bacteria and identification of the isolated strain
As described before, the strain QD-5 was elected for the further analysis. Analysis of the 16S rDNA on sequence similarity revealed that the closest relative of the strain QD-5 was Vibrio alginolyticus strain W047 (JF836178.1) (99%). In the phylogenetic tree, the strain QD–5 was grouped within the genus Vibrio (Fig. 1). Consequently, the strain QD-5 was designated as Vibrio sp. QD-5 and it has been deposited at the China General Microbiological Culture Collection Center (CGMCC) under accession number 14172.
Figure
1.
Phylogenetic analysis of strain QD-5. Phylogenetic tree was constructed through the neighbor-joining algorithm using MEGA6.0, and the evolutionary distance was estimated by the maximum composite likelihood (MCL) method (Tamura et al., 2011).
3.2
Information about the aly–IV gene and the deduced amino acid sequence
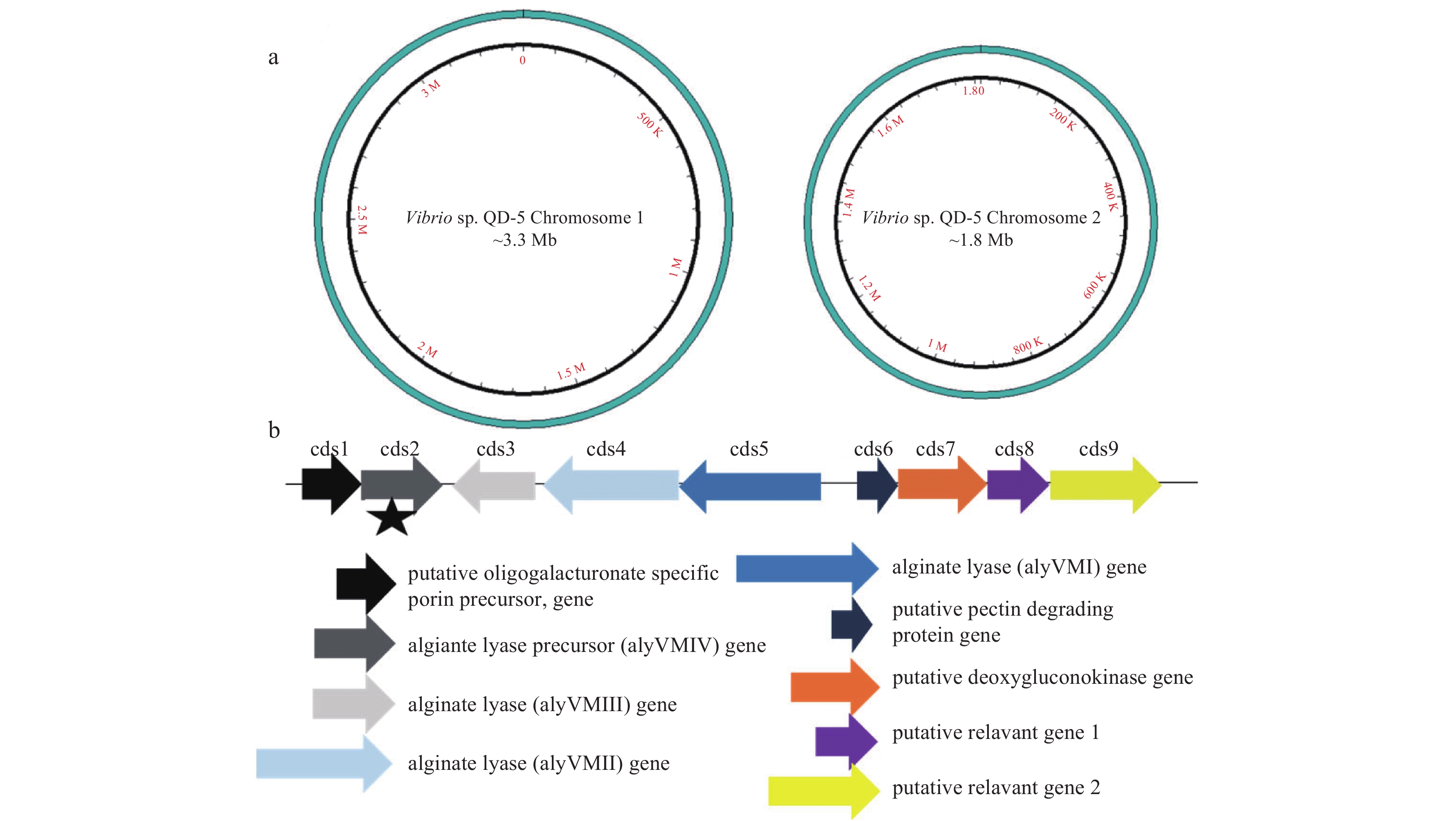
Using the sequenced genomic data in this work, the draft genome was further refined to contain a total genome size of 1 118 293 600 bp with an average GC content of 44.75% and two circular plasmids sized at approximately 3.3 Mbp and 1.8 Mbp (Fig. 2a). Annotation of the genome showed that an open reading frame coding for polysaccharide depolymerases located on the chromosome 2 (Fig. 2b). One of the genes, aly–IV, was cloned and sequenced. It consisted of 1 560 bp and the open reading frame (ORF) encoded 520 amino acids without signal peptide (Fig. 3). It was subjected for BLAST analysis in NCBI database. Result showed that the amino acid sequence display high similarity with several alginate lyase (WP_017821221.1, WP_054574336.1, WP_054578990.1, AIY22655.1, WP_017634226.1, WP_053307759.1, WP_053349966.1, WP_005388518.1, 99% of sequence identity). But these alginate lyases have never been characterized. The mature protein had a calculated molecular mass of 61.92 kDa and a pI value of 5.12. Sequence alignments showed that, Aly–IV was assigned to polysaccharide lyase family 7 (PL7). Conserved domain analysis of the putative protein revealed that the protein contained two domains, including F5/8 type C domain (pfam00754) and Alginate lyase2 domain (pfam08787) (Fig. 3). Alginate lyases in PL7 contain three highly conserved motifs, (R/E)(S/T/N)ELR, Q(I/V)H and YFKAG(I/V)YNQ, which composed the active center and were crucial for substrate recognition and catalysis (Yamasaki et al., 2005). Sequence alignment of Aly-IV with other PL7 alginate lyases indicated that the conserved motifs in Aly–IV were RTELR, YFKAGVYNQ and QIH (Fig.4). It has been confirmed that the isoleucine residue in QIH motif is essential for recognition of the polyG or G–G bond (Deng et al., 2014).
Figure
2.
The genome and alginate lyase gene cluster of the Vibrio sp. QD-5. a. The draft genome of the strain Vibrio sp. QD-5. b. Four alginate lyase genes in the genome of the Vibrio sp. QD-5, they were cds2, cds3, cds4, cds5. The cds1, cds6, cds7, cds8, cds9 were the genes relevant to the alginate lysae gene. The asterisk is the position of aly-IV.
Figure
3.
Sequence analysis of Aly-IV. a. Functional annotation of the predicted protein. b. Nucleotide and deduced amino acid sequence of the Aly-IV. The signal peptide was underlined straight line. Dashed line was the F5/8 type C domain. The dotted line was Alginate lyase2 domain.
Figure
4.
Amino acid alignment of Aly-IV with other PL7 alginate lyase. BAV53312.1, an alginate lyase from Vibrio algivorus; BAV10560.1, an endo-type alginate lyase from Falsirhodobacter sp. alg1; ANB32492.1, an alginate lyase from Halomonas sp. Victoria JH; BAJ62034.1, an alginate lyase from Microbulbifer sp. 6532A; ACM89454.1, an alginate lyase from Pseudoalteromonas sp. CY24; BAK40155.1, an alginate lyase from Saccharophagus sp. Myt-1; ADW41663.1, an alginate lyase precursor from Vibrio midae; BAH79133.1, an alginate lyase from Vibrio sp. A9m; ALP75563.1, an alginate lyase from Vibrio sp. BZM-1; and ALP75562.1, an alginate lyase from Vibrio litoralis. Similar amino acid residues among the alginate lyases were shaded in gray. Conserved regions were boxed. Arrows represented the conserved sequences below them.
3.3
Purification of the recombination enzyme Aly–IV
To characterize the recombinant enzyme, the gene aly–IV was cloned into an expression vector pET–22b (+) and expressed in E. coli BL21 (DE3). Tests showed that the recombinant enzyme, Aly–IV, was expressed in the form of IB. To obtain the active enzyme Aly–IV, the IB was denatured by sarkosyl and refolded via dilution. A large amount of refolded enzyme were obtained and purified by nickel affinity chromatography followed by gel filtration chromatography. The apparent molecular weight was 62 kDa on the SDS–PAGE (Fig. 5). The final activity of the enzyme reached 1 256.78 U/mg, which was different from the alginate lyase from other Vibrio sp. The activity of Alyb from Vibrio sp. W13 reached 457 U/mg (Zhu et al., 2015). While AlyVI from Vibrio sp. QY101 possessed the activity of 6 380 U/mg (Han et al., 2004), the activities of AlyAL–28 and AlyATCC only reached 1.89 and 1.93 U/mg, respectively (Kitamikado et al., 1992).
Figure
5.
SDS-PAGE analysis of the purified enzyme Aly-IV. Proteins were electrophoresed on 12.5% acrylamide gel and stained with Coomassie Brilliant Blue G-250. Lane 1: 10 μL of the protein (150 μg/mL) purified by Ni-NTA affinity chromatography. Lane M: protein mass markers. Lane 2: 20 μL of the protein (10 μg/mL) purified by Superdex-200 Increase10/300.
3.4
Characteristics of the recombinant enzyme Aly-IV
The optimal temperature of the enzyme Aly–IV was 35°C (Fig. 6a) and remained stable when incubated at temperatures below 30°C for 30 min (Fig. 6b). Aly–IV showed the highest activity in the Tris–HCl buffer at pH of 8.9 (Fig. 6c). Compared with other alginate lyase from Vibrio strains, such as AlySY08 with the optimal pH of 7.6 (Li et al., 2017), AlyVI with the optimal pH of 7.5 (Han et al., 2004), Algb with the optimal pH of 8.0 (Zhu et al., 2015), AlyAL-128 with the optimal pH of 7.8 (Kitamikado et al., 1992), AlyATCC with the optimal pH of 8.2 (Kitamikado et al., 1992), the alginate lyase Aly-IV had the higher pH of 8.9.
Figure
6.
Biochemical characterization of the recombinant alginate lyase Aly-IV. a. Reaction temperature profile of Aly-IV. The activity was assayed at pH 8.9. b. Thermal stability of Aly-IV. The enzyme was incubated at 10–55°C for 30 min, and then the residual enzyme activity was measured at 35°C. c. Reaction pH profile of Aly-IV. The activity was assayed at 35°C in different buffers as described before. d. pH stability of Aly-IV. The residual activity was determined by measuring the enzyme activity at 35°C after incubation at various pHs for 3 h in ice-bath. The data represent the mean of three experimental repeats with SD≤5%.
The pH stability test of the Aly–IV was conducted in ice–bath for 3 h (Fig. 6d). The pH stability of the enzyme in different buffer was related to the type of the buffer. No more than 25% of Aly–IV activity was retained in the Na2HPO4–citric acid buffer (pH 5.5–6.0). In the PB buffer (pH 5.8–8.0), the residual activity was lower than 70% of its initial activity. In the Tris–HCl buffer (pH 7.0–8.9), the activity retained more than 50%. Notably, in the Tris–HCl buffer with pH 7.0, the enzyme activity was almost unaffected. In the glycine–NaOH buffer (pH 9.0–10.6), the residual activity was from 42% to 90%.
The enzyme Aly–IV activity varied in different buffers at the same pH, suggesting that the buffer ions affect the enzyme activity. Therefore, the effect of metal ions on the enzyme activity was examined. The effects of the NaCl on the activity of the enzyme Aly–IV showed in Fig. 7a. The enzyme activity was inhibited when the concentration was more than 25 mmol/L. Moreover, the activity of Aly–IV was strongly inhibited by 1 mmol/L Ba2+, Al3+, Ni2+, Zn2+, and Pb2+, but promoted by 1 mmol/L Ca2+ (Fig. 7b), increased with K+ and Mg2+ at the concentration of 5 and 10 mmol/L. The effects of mental ions on Aly-IV were similar to that on Algb (Zhu et al., 2015) and AlyAL-128 (Kitamikado et al., 1992). Alginate lyases from different sources were affected by different mental ions. This may be due to the differences of their three-dimensional structures.
Figure
7.
The effects of NaCl concentration and metal ions on the activity of Aly-IV. a. Effect of NaCl concentration on aly-SJ02 activity. The activity was measured at 35°C in Tris-HCl buffer containing the indicated NaCl concentrations. The activity in 0 mol/L NaCl was taken as 100%. b. Metal ions effects on the activity of Aly-IV. The enzyme activity was measured at 35°C in Tris–HCl buffer containing different metal ions. The activity of aly-SJ02 without any metal ion or inhibitor in the reaction mixture was taken as control (100%). The data represent the mean of three experimental repeats with SD≤5%.
3.5
Enzymtic hydrolysis of sodium alginate by Aly–IV
To study degradation characteristics of Aly–IV, the amount of degradation product was measured at different time points. As shown in Fig. 8, the amount of product increased considerably from 0.5 to 6 h, and plateaued after 6 h, due to the loss of the enzyme activity when incubated at 35°C for a long time.
Figure
8.
Enzymatic hydrolysis of sodium alginate by Aly-IV. The course of hydrolytic procedure was monitored by measuring the amount of product during the hydrolysis.
3.6
Substrate specificity of the recombinant enzyme
To investigate the substrate specificity, sodium alginate, polyG, polyM and polyGM were used as the substrates for the recombinant enzyme Aly–IV. In contrast to polyG, the activity of Aly–IV towards polyM was negligible. This illustrated that Aly–IV broke G–G links specifically. The observed lower activity towards polyM and polyGM was speculated to be caused by purity of the substrates.
The Km values of the enzyme towards sodium alginate, polyG, polyM and polyGM were 0.22, 0.33, 2.89 and 1.42 mg/mL, respectively (Table 1). The lower Km values of the enzyme with sodium alginate and polyG than that with polyM and polyGM suggested that the enzyme had the higher affinity to sodium alginate and polyG. As previously reported, most of the alginate lyases were polyM-preferred alginate lyases. Only few polyG-preferred alginate lyases were discovered, such as AlySY08 from marine bacterium Vibrio sp. SY08 (Li et al., 2017), AlyV5 from Vibrio sp. QY105 (Wang et al., 2013), and AlyAL-128 from Vibrio harveyi AL-128 (Kitamikado et al., 1992).
3.7
Reaction products and modes of action
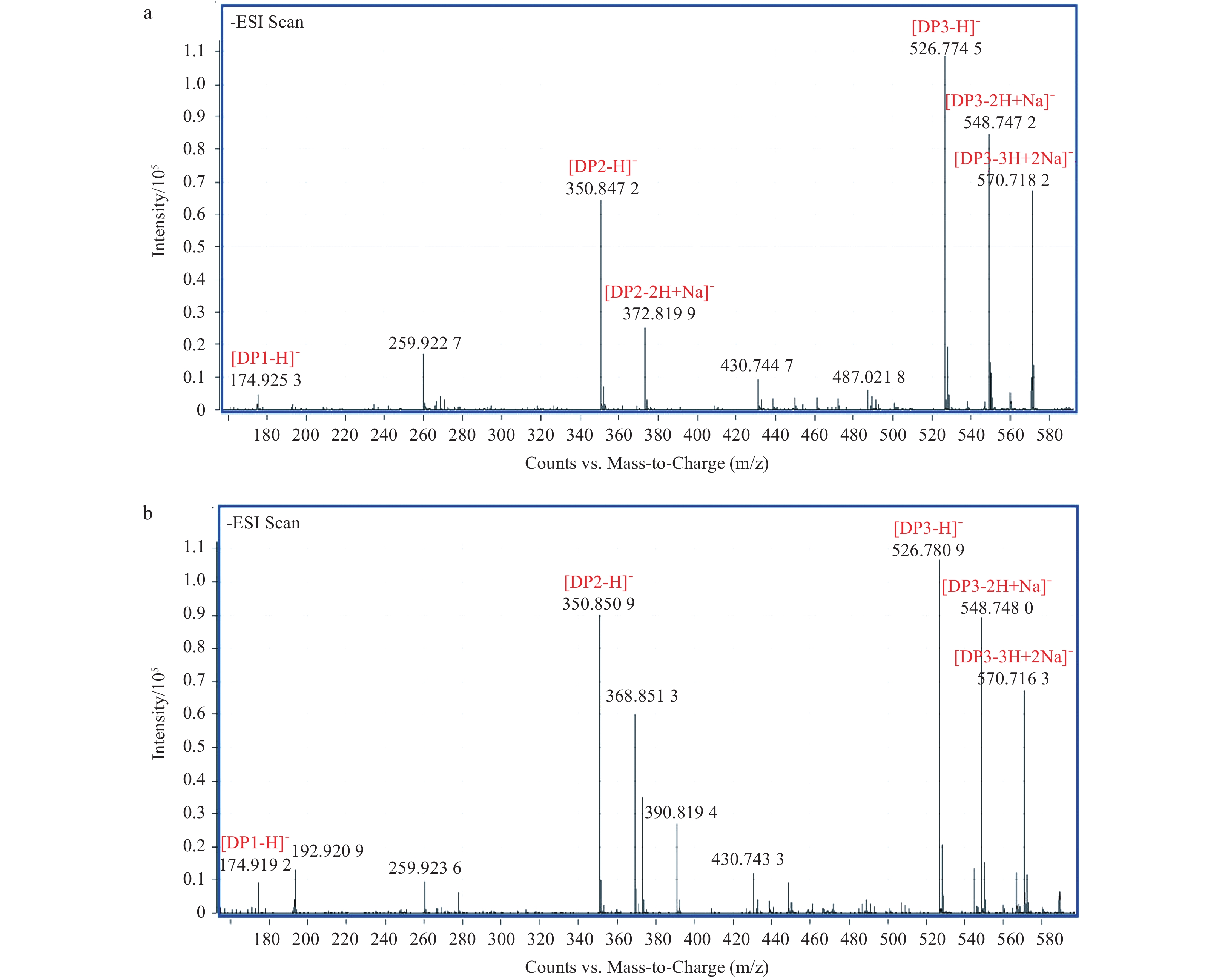
To analyze the degradation products of the recombinant enzyme Aly–IV, 5 μL samples taken at different time points were subjected to TLC assy. The distributions of the products from the substrates of sodium alginate and polyG were similar (Figs 9a, b). The products of polyM and polyGM were not detected on the TLC plates (Figs 9c, d). The reaction products of sodium alginate and polyG were further identified by negative ESI–TOF–MS. As shown in Fig. 10, the degradation products of sodium alginate and polyG mainly consist of DP1 and DP2. The enzyme had no activity to polyM and polyGM, so the DP of the products from polyM and polyGM was not identified by negative ESI–TOF–MS.
Figure
9.
TLC analysis of the degradation products of the recombinant enzyme towards sodium alginate (a), polyG (b), polyM (c) and polyMG (d). Samples were taken out at intervals. Line M: galacturonic acid. The solvent system was 1-butanol/ethanol/water 2:1:1 and products were visualized by heated the plate at 85°C for 10 min after spray with a diphenylamine/aniline/phosphate reagent.
Figure
10.
ESI–TOF-MS analyses of the end products of Aly–IV. The samples applied to ESI–TOF–MS was reacted for 6 h. a. The degradation products of sodium alginate and b. the degradation products of polyG.
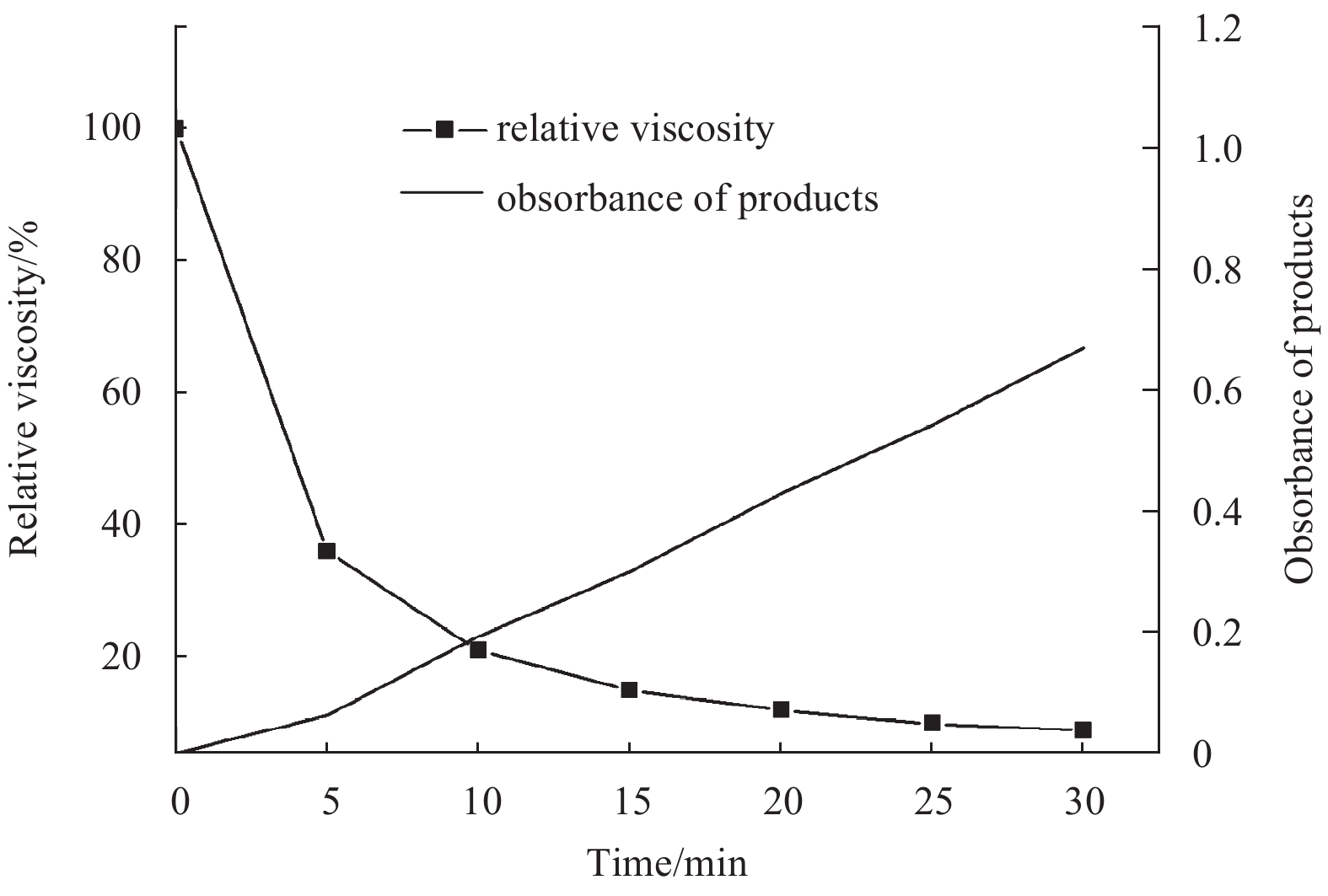
The action mode of the recombinant enzyme Aly–IV was monitored in the viscometric assay. The viscosity of the alginate solution rapidly decreased in the first 5 min, whereas the values of absorbance of reducing products increased very slowly (Fig. 11). The distribution of the hydrolysate on the TLC plate suggested that the enzyme Aly–IV acts in an endo-lytic mode. The endo-lytic mode actions of enzyme Alm from Agarivorans sp. JAM-A1m (Kobayashi et al., 2009) and enzyme AlySY08 from Vibrio sp. SY08 (Li et al., 2017) were confirmed in the same method.
Figure
11.
Viscosity reduction during enzymatic degradation of alginate. The initial viscosity of the reaction mixture was taken as 100%. Square filled with dotted line is rate of viscosity reduction; triangle filled with solid line is absorbance of reducing products.
In this study, an alginate lyase gene, aly-IV, was cloned and expressed in E. coil BL21. The purification and characterization of this enzyme were reported for the first time. Its optimal temperature and pH was 35°C and 8.9, respectively. Like other alginate lyases, such as AL-128 from Vibrio harveyi (Kitamikado et al., 1992), rA9mT from Vibrio sp. JAM-A9m (Uchimura et al., 2010), AlYSY08 from Vibrio sp. SY08 (Li et al., 2017), Aly-IV was also a salt-activated enzyme whose activity was increased three times at 15 mmol/L NaCl. The enzyme Aly-IV may be activated by K+, Mg2+ and Ca2+, which was similar to other polyG-specific alginate lyase.
As shown in Fig. 4, PL7 alginate lyases contain three highly conserved amino acid sequences, (S/T/N)EL, Q(I/V)H, and YFKAG(V/I)YNQ, which were thought to play an important role in catalytic activity or the folding of the structure (Zhu et al., 2015). Researches revealed that enzymes containing QIH or QVH in the conserved region had substrate specificity of G-block and M-block, respectively (Yamasaki et al., 2005). However, Aly-IV seems to prefer poly-G, like ALY-1 from Corynebacterium sp. ALY-1 (Matsubara et al., 1995), A1-I from Sphingomonas sp. A1 (Miyake et al., 2003) and A1m from Agarivorans sp. JAM-A1m (Kobayashi et al., 2009).
Alginate extracted from seaweed has a wide range of applications. A lot of organisms are also able to exploit M and G units from depolymerized alginates as carbon and energy sources. Most of them can secrete the alginate lyase to degrade alginate. So far, hundreds of alginate lyases were found from a variety of organisms, such as marine algae, marine mollusks (Littorina spp., Haliotis spp., Turbo cornutus.), and a wide range of marine and terrestrial bacteria. Alginate lyases belong to the member of the polysaccharide lyases. But the concern is: Why so many organisms produce alginate lyase, including even the bacteria from deep-sea (Wong et al., 2000)? Is not the alginate everywhere? Why the gene sequences of alginate lyases from the different sources are so diverse (Uchimura et al., 2010)? So far, these questions remained unanswered. We speculate that alginate lyase may not only degrade alginate, but also degrade other polysaccharides as alginate lyases in organisms have multiple substrate specificities. Although no report has been seen yet, we will attempt to confirm this speculation in future.
Acknowledgements
We thank Shuyan Wang and Jiaqiang Wei for the sample collection, Shuqi Wu for the bioinformatics analysis and Hao Chen for the study design and the manuscript review.
Aarstad O A, Tøndervik A, Sletta H, et al. 2012. Alginate sequencing: an analysis of block distribution in alginates using specific alginate degrading enzymes. Biomacromolecules, 13(1): 106–116. doi: 10.1021/bm2013026
An Q D, Zhang G L, Wu H T, et al. 2009. Alginate-deriving oligosaccharide production by alginase from newly isolated Flavobacterium sp. LXA and its potential application in protection against pathogens. Journal of Applied Microbiology, 106(1): 161–170. doi: 10.1111/jam.2008.106.issue-1
Atala A, Kim W, Paige K T, et al. 1994. Endoscopic treatment of vesicoureteral reflux with a chondrocyte-alginate suspension. The Journal of Urology, 152(2 Pt 2): 641–643
Boyd J, Turvey J R. 1978. Structural studies of alginic acid, using a bacterial poly-α-l-guluronate lyase. Carbohydrate Research, 66(1): 187–194. doi: 10.1016/S0008-6215(00)83251-6
Butler D M, Østgaard K, Boyen C, et al. 1989. Isolation conditions for high yields of protoplasts from Laminaria Saccharina and L. Digitata (Phaeophyceae). Journal of Experimental Botany, 40(11): 1237–1246. doi: 10.1093/jxb/40.11.1237
Deng Shuping, Ye Jiang, Xu Qingqing, et al. 2014. Structural and functional studies on three alginate lyases from Vibrio alginolyticus. Protein and Peptide Letters, 21(2): 179–187
Fremond B, Malandain C, Guyomard C, et al. 1993. Correction of bilirubin conjugation in the Gunn Rat using hepatocytes immobilized in alginate gel beads as an extracorporeal bioartificial liver. Cell Transplant, 2(6): 453–460. doi: 10.1177/096368979300200603
Gacesa P. 1992. Enzymic degradation of alginates. International Journal of Biochemistry, 24(4): 545–552. doi: 10.1016/0020-711X(92)90325-U
Han Feng, Gong Qianhong, Song Kai, et al. 2004. Cloning, sequence analysis and expression of gene alyVI encoding alginate lyase from marine bacterium Vibrio sp. QY101. DNA Sequence, 15(5–6): 344–350. doi: 10.1080/10425170400019300
Hou Baobing, Liu Shulai, Zhang Jianyou, et al. 2009. Optimization of alginate-degrading bacterial fermentation process for alginate lyase production. Fisheries Science (in Chinese), 28(11): 667–670
Hu Xiaoke, Jiang Xiaolu, Guan Huashi. 2003. Isolation of protoplasts from undaria pinnatifida by alginate lyase digestion. Journal of Ocean University of China, 2(1): 58–61
Hu Xiaoke, Jiang Xiaolu, Hwang H, et al. 2004. Promotive effects of alginate-derived oligosaccharide on maize seed germination. Journal of Applied Phycology, 16(1): 73–76. doi: 10.1023/B:JAPH.0000019139.35046.0c
Huang Lishuxin, Zhou Jungang, Li Xiao, et al. 2013. Characterization of a new alginate lyase from newly isolated Flavobacterium sp. S20. Journal of Industrial Microbiology and Biotechnology, 40(1): 113–122. doi: 10.1007/s10295-012-1210-1
Inoue A, Mashino C, Kodama T, et al. 2011. Protoplast preparation from laminaria japonica with recombinant alginate lyase and cellulase. Marine Biotechnology, 13(2): 256–263. doi: 10.1007/s10126-010-9290-2
Iwamoto M, Kurachi M, Nakashima T, et al. 2005. Structure-activity relationship of alginate oligosaccharides in the induction of cytokine production from RAW264. 7 cells . FEBS Letters, 579(20): 4423–4429. doi: 10.1016/j.febslet.2005.07.007
Kitamikado M, Tseng C H, Yamaguchi K, et al. 1992. Two types of bacterial alginate lyases. Applied and Environmental Microbiology, 58(8): 2474–2478
Kobayashi T, Uchimura K, Miyazaki M, et al. 2009. A new high-alkaline alginate lyase from a deep-sea bacterium Agarivorans sp. Extremophiles, 13(1): 121–129. doi: 10.1007/s00792-008-0201-7
Li Liyan, Jiang Xiaolu, Guan Huashi, et al. 2011. Preparation, purification and characterization of alginate oligosaccharides degraded by alginate lyase from Pseudomonas sp. HZJ 216. Carbohydrate Research, 346(6): 794–800. doi: 10.1016/j.carres.2011.01.023
Li Shangyong, Wang Linna, Hao Jianhua, et al. 2017. Purification and characterization of a new alginate lyase from marine bacterium Vibrio sp. SY08. Marine Drugs, 15(1): E1
Matsubara Y, Iwasaki K, Muramatsu T. 1995. Purification and Characterization of Extracellular Alginate Lyase from Corynebacterium sp. ALY-1 Strain. Japan: The Society for Biotechnology
Miyake O, Hashimoto W, Murata K. 2003. An exotype alginate lyase in Sphingomonas sp. A1: overexpression in Escherichia coli, purification, and characterization of alginate lyase IV (A1-IV). Protein Expression and Purification, 29(1): 33–41. doi: 10.1016/S1046-5928(03)00018-4
Østgaard K. 1993. Determination of alginate composition by a simple enzymatic assay. Hydrobiologia, 260(1): 513–520
Steinlechner-Maran R, Eberl T, Kunc M, et al. 1996. Oxygen dependence of respiration in coupled and uncoupled endothelial cells. American Journal of Physiology, 271(6 Pt 1): C2053–C2061
Tamura K, Peterson D, Peterson N, et al. 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution, 28(10): 2731–2739. doi: 10.1093/molbev/msr121
Tao Hu, Liu Wenjun, Simmons B N, et al. 2010. Purifying natively folded proteins from inclusion bodies using sarkosyl, triton X-100, and CHAPS. Biotechniques, 48(1): 61–64. doi: 10.2144/000113304
Uchimura K, Miyazaki M, Nogi Y, et al. 2010. Cloning and sequencing of alginate lyase genes from deep-sea strains of Vibrio and Agarivorans and characterization of a new Vibrio enzyme. Marine Biotechnology (NY), 12(5): 526–533. doi: 10.1007/s10126-009-9237-7
Wang Ya, Guo Enwen, Yu Wengong, et al. 2013. Purification and characterization of a new alginate lyase from a marine bacterium Vibrio sp. Biotechnology Letters, 35(5): 703–708. doi: 10.1007/s10529-012-1134-x
Wong T Y, Preston L A, Schiller N L. 2000. Alginate lyase: review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annual Review of Microbiology, 54: 289–340. doi: 10.1146/annurev.micro.54.1.289
Yamasaki M, Ogura K, Hashimoto W. 2005. A structural basis for depolymerization of alginate by polysaccharide lyase family-7. Journal of Molecular Biology, 352(1): 11–21. doi: 10.1016/j.jmb.2005.06.075
Zhang Ruxue, Zhou Jinhuang, Jia Zhengping, et al. 2004. Hypoglycemic effect of Rehmannia glutinosa oligosaccharide in hyperglycemic and alloxan-induced diabetic rats and its mechanism. Journal of Ethnopharmacology, 90(1): 39–43. doi: 10.1016/j.jep.2003.09.018
Zhu Benwei, Tan Haidong, Qin Yuqi, et al. 2015. Characterization of a new endo-type alginate lyase from Vibrio sp. W13. International Journal of Biological Macromolecules, 75: 330–337. doi: 10.1016/j.ijbiomac.2015.01.053
Zong Wanrong, Cheong K L, Wu Dingtao, et al. 2014. Preparation and purification of raffinose family oligosaccharides from Rehmannia glutinosa Libosch. by fast protein liquid chromatography coupled with refractive index detection. Separation and Purification Technology, 138: 98–103. doi: 10.1016/j.seppur.2014.10.001
Fen Yan, Junying Chen, Ting Cai, et al. Cloning, expression, and characterization of a novel endo‐type alginate lyase from Microbulbifer sp. BY17. Journal of the Science of Food and Agriculture, 2022, 102(11): 4522. doi:10.1002/jsfa.11807
2.
Li Sha, Minghai Huang, Xiaonan Huang, et al. Cloning and Characterization of a Novel Endo-Type Metal-Independent Alginate Lyase from the Marine Bacteria Vibrio sp. Ni1. Marine Drugs, 2022, 20(8): 479. doi:10.3390/md20080479
3.
Shu-Kun Gao, Rui Yin, Xiao-Chen Wang, et al. Structure Characteristics, Biochemical Properties, and Pharmaceutical Applications of Alginate Lyases. Marine Drugs, 2021, 19(11): 628. doi:10.3390/md19110628
4.
Wenwen Cheng, Xuanyu Yan, Jiali Xiao, et al. Isolation, identification, and whole genome sequence analysis of the alginate-degrading bacterium Cobetia sp. cqz5-12. Scientific Reports, 2020, 10(1) doi:10.1038/s41598-020-67921-7
Table
1.
Specific activity and kinetic parameters of the recombinant enzyme Aly-IV towards sodium alginate, polyG, polyM and polyGM
Substrate
Km/mg∙mL–1
Vmax/U∙mg–1
Relative activity/%
Sodium alginate
0.22
1 056.8
100
polyG
0.33
1 247.6
68.41
polyM
2.89
506.3
0.53
polyGM
1.42
711.8
1.23
Note: The data were the means of three experimental repeats with SD<0.05. The initial reaction velocity was defined as the increasing absorbance at 235 nm of the products produced by recombinant enzyme Aly-IV at the first 5 min.
Figure 1. Phylogenetic analysis of strain QD-5. Phylogenetic tree was constructed through the neighbor-joining algorithm using MEGA6.0, and the evolutionary distance was estimated by the maximum composite likelihood (MCL) method (Tamura et al., 2011).
Figure 2. The genome and alginate lyase gene cluster of the Vibrio sp. QD-5. a. The draft genome of the strain Vibrio sp. QD-5. b. Four alginate lyase genes in the genome of the Vibrio sp. QD-5, they were cds2, cds3, cds4, cds5. The cds1, cds6, cds7, cds8, cds9 were the genes relevant to the alginate lysae gene. The asterisk is the position of aly-IV.
Figure 3. Sequence analysis of Aly-IV. a. Functional annotation of the predicted protein. b. Nucleotide and deduced amino acid sequence of the Aly-IV. The signal peptide was underlined straight line. Dashed line was the F5/8 type C domain. The dotted line was Alginate lyase2 domain.
Figure 4. Amino acid alignment of Aly-IV with other PL7 alginate lyase. BAV53312.1, an alginate lyase from Vibrio algivorus; BAV10560.1, an endo-type alginate lyase from Falsirhodobacter sp. alg1; ANB32492.1, an alginate lyase from Halomonas sp. Victoria JH; BAJ62034.1, an alginate lyase from Microbulbifer sp. 6532A; ACM89454.1, an alginate lyase from Pseudoalteromonas sp. CY24; BAK40155.1, an alginate lyase from Saccharophagus sp. Myt-1; ADW41663.1, an alginate lyase precursor from Vibrio midae; BAH79133.1, an alginate lyase from Vibrio sp. A9m; ALP75563.1, an alginate lyase from Vibrio sp. BZM-1; and ALP75562.1, an alginate lyase from Vibrio litoralis. Similar amino acid residues among the alginate lyases were shaded in gray. Conserved regions were boxed. Arrows represented the conserved sequences below them.
Figure 5. SDS-PAGE analysis of the purified enzyme Aly-IV. Proteins were electrophoresed on 12.5% acrylamide gel and stained with Coomassie Brilliant Blue G-250. Lane 1: 10 μL of the protein (150 μg/mL) purified by Ni-NTA affinity chromatography. Lane M: protein mass markers. Lane 2: 20 μL of the protein (10 μg/mL) purified by Superdex-200 Increase10/300.
Figure 6. Biochemical characterization of the recombinant alginate lyase Aly-IV. a. Reaction temperature profile of Aly-IV. The activity was assayed at pH 8.9. b. Thermal stability of Aly-IV. The enzyme was incubated at 10–55°C for 30 min, and then the residual enzyme activity was measured at 35°C. c. Reaction pH profile of Aly-IV. The activity was assayed at 35°C in different buffers as described before. d. pH stability of Aly-IV. The residual activity was determined by measuring the enzyme activity at 35°C after incubation at various pHs for 3 h in ice-bath. The data represent the mean of three experimental repeats with SD≤5%.
Figure 7. The effects of NaCl concentration and metal ions on the activity of Aly-IV. a. Effect of NaCl concentration on aly-SJ02 activity. The activity was measured at 35°C in Tris-HCl buffer containing the indicated NaCl concentrations. The activity in 0 mol/L NaCl was taken as 100%. b. Metal ions effects on the activity of Aly-IV. The enzyme activity was measured at 35°C in Tris–HCl buffer containing different metal ions. The activity of aly-SJ02 without any metal ion or inhibitor in the reaction mixture was taken as control (100%). The data represent the mean of three experimental repeats with SD≤5%.
Figure 8. Enzymatic hydrolysis of sodium alginate by Aly-IV. The course of hydrolytic procedure was monitored by measuring the amount of product during the hydrolysis.
Figure 9. TLC analysis of the degradation products of the recombinant enzyme towards sodium alginate (a), polyG (b), polyM (c) and polyMG (d). Samples were taken out at intervals. Line M: galacturonic acid. The solvent system was 1-butanol/ethanol/water 2:1:1 and products were visualized by heated the plate at 85°C for 10 min after spray with a diphenylamine/aniline/phosphate reagent.
Figure 10. ESI–TOF-MS analyses of the end products of Aly–IV. The samples applied to ESI–TOF–MS was reacted for 6 h. a. The degradation products of sodium alginate and b. the degradation products of polyG.
Figure 11. Viscosity reduction during enzymatic degradation of alginate. The initial viscosity of the reaction mixture was taken as 100%. Square filled with dotted line is rate of viscosity reduction; triangle filled with solid line is absorbance of reducing products.



 DownLoad:
DownLoad:

















 DownLoad:
DownLoad: