

Morphological, phylogenetic and metabolite profile of Prorocentrum clipeus, a newly recorded epiphytic dinoflagellate in the northern Yellow Sea
-
Abstract: More than 30 species of benthic Prorocentrum have been identified, some of which produce okadaic acid (OA) and its derivatives, dinophysistoxins (DTXs), which cause diarrhetic shellfish poisoning (DSP). Increasing numbers of benthic Prorocentrum species have been reported in tropical and subtropical waters of China. In contrast, only a few benthic Prorocentrum species have been reported in temperate waters. In this study, morphological descriptions obtained using light microscopy, scanning electron microscopy and molecular characterization of one Prorocentrum clipeus strain isolated from the Yellow Sea are presented. Prorocentrum clipeus cells were nearly circular in shape, with a collar, ridge, and one protrusion. The periflagellar area was wide U-shaped, with two curved projections on platelet 1a. Nine periflagellar platelets of different sizes were observed. The morphology closely fits that of the species isolated from other locations. Phylogenetic analysis based on the molecular sequences of the small subunit (SSU) rDNA, internal transcribed spacer (ITS), and large subunit (LSU) rDNA was performed. A comprehensive metabolomic analysis incorporating target, suspect and non-target screenings was first applied to investigate the intracellular and extracellular metabolite profiles of the current isolate of P. clipeus. According to the results of the target and suspect screenings, 179 metabolites or toxins produced by DSP-related algal species, including OA, dinophysistoxin-1 (DTX1), dinophysistoxin-2 (DTX2) and pectenotoxin-2 (PTX2), were not detected. Non-target screening involving feature-based molecular networking (FBMN) provided a global view of major metabolites produced by the P. clipeus DF128 strain and revealed 23 clusters belonging to at least 13 compound classes, with organometallic compounds, lipids and lipid-like molecules, phenylpropanoids and polyketides, and benzenoids as major types. To date, this is the first record of the characterization of P. clipeus in samples from Chinese waters. Our results support the wide distribution of epibenthic Prorocentrum species.
-
Key words:
- Prorocentrum /
- taxonomy /
- phylogeny /
- metabolite profiling /
- Yellow Sea
-
1. Introduction
The dissolved oxygen (DO) in seawater is key to shaping the growth and reproduction of marine organisms. It is an important contributor to the biogeochemical cycle. Overall, the magnitude of the DO concentration reflects the health of an aquatic system. Hypoxia occurs when the rate of oxygen consumption in seawater exceeds the rate at which it is replaced. When the DO concentration in seawater is as low as 3–4 mg/L, many marine organisms are adversely affected (Diaz, 2001; Wu, 2002; Vaquer-Sunyer and Duarte, 2008; Zhai et al., 2012). At DO concentration values of less than 3 mg/L, certain fish begin to die, and the number of benthic organisms is greatly reduced. When the DO is completely consumed, i.e., oxygen-free (no oxygen), which results in anoxia, most marine life cannot survive (Li et al., 2002; Zhang et al., 2010; Zhai et al., 2012). Therefore, selecting different hypoxia thresholds in different regions is beneficial to accurately assessing the occurrence of hypoxic events.
Biochemical processes involving oxygen consumption and water stratification (e.g., density differences) hinder the exchange between water surface with high levels of oxygen and hypoxic bottom water, which directly causes hypoxia (Turner et al., 1987; Rabalais et al., 2002; Hetland and Dimarco, 2008). Taft et al. (1980) pointed out that fresh water increased density stratification in Chesapeake Bay in spring, resulting in hypoxia of the water below a depth of 10 m. In addition, studies have also shown that hypoxic areas such as Chesapeake Bay (Hagy et al., 2004) and the middle of the Baltic Sea (Conley et al., 2002) are related to the submarine topography. Rabalais et al. (2007, 2010) found that the Mississippi River Estuary injected a large amount of fresh water and excess nutrient salts into the northern part of the Gulf of Mexico. This large amount of hot fresh water promoted the formation of high-density stratified structures in summer, and the input of excess nutrient salts led to intensification of the water eutrophication. The combination of these two processes induced the formation of seasonal hypoxia at the bottom. The hypoxia at the bottom of the Changjiang River Estuary in summer is caused by the large quantity of nutrients carried by diluted water, leading to stratification and a marked increases in primary productivity (Li et al., 2002; Luo et al., 2005; Zhang et al., 2007; Wei et al., 2007; Chen et al., 2007). The degree of summer hypoxia is correlated with the magnitude of the river flux in estuary/inshore areas that are controlled by runoff water. Increased nutrient delivery can increase the primary productivity and lead to oxygen consumption, but it can also decrease the residence time and increase flushing, reducing the duration of hypoxic events, such as those in the Chesapeake Bay, Neuse, and Pamlico estuaries in the United States and in the Black Sea, Adriatic Sea, German Gulf, and North Sea in Europe (Diaz, 2001; Lin et al., 2006, 2008; Hagy et al., 2004).
The Bohai Sea (BS) is a semi-enclosed sea connected to the Yellow Sea through an opening east of the Bohai Strait (Fig. 1). It is very shallow and has a mean depth of 18 m. The general circulation pattern of the BS is shaped by seawater, which enters north of the Bohai Strait and exits in the south. This trend results in the entire BS having an anticlockwise circulation (Huang et al., 1999; Wang et al., 2010; Ding et al., 2019; Wu et al., 2019). Many rivers, such as the Huanghe and Haihe rivers, converge in the BS area. These rivers carry large amounts of organic materia from the inland region, making the BS a natural fishing ground that is rich in seafood such as shrimp, crab, and yellow croaker. The spawning and breeding periods of economically important fish, shrimp, and crab in the BS are between March and August. The deep-water area in the central BS (CBS) is the distribution center for the migration of organisms of dietary importance. However, it acts as their wintering ground. With the rapid economic development and urbanization of the Bohai Rim region, the flux of land-based pollutants into the sea has increased. According to the 2015 Marine Environment Bulletin of the Beihai Region, from 2001 to 2015, the coastal waters of the BS became increasingly polluted, and this pollution manifested as the frequent occurrence of red tides. The waters that underwent eutrophication were mainly located in the top of Liaodong Bay, Bohai Bay, and Laizhou Bay. Shellfish tend to be plagued by frequent red tides due to increased eutrophication in these three major bays of the BS (Bulletin of the State of the Marine Environment of China, 2010). For example, in the summer of 1997 and 1998, many farmed scallops died in the coastal waters around the BS (Zhai et al., 2012), most likely due to hypoxia. Hypoxic events in the CBS often occur near double-centered cold water masses (DCCWM). Wei et al. (2021) analyzed data for August acquired from a station near the cold water mass near the northern BS using the National Oceanic Standard Profile Measurement dataset (1978–2018). They reported that severe oxygen depletion occurred during the summer from 2006 to 2018. Zhai et al. (2012) investigated DO concentration and pH data from 20–23 stations in and around the CBS for June and August 2011. They found that the DO concentration of the bottom water decreased significantly and that the water was acidified in a zonal area at depths of 20–35 m in the northwest and northern coastal regions of the BS (red dashed boxes in Fig. 1a). Zhang et al. (2016) conducted a summer survey of the BS in 2014. They reported that the formation of a seasonal thermocline in summer was the main physical process in the hypoxic zone at the bottom, and the oxygen consumption of mineralization and decomposition processes was also the reason for the formation of a hypoxic zone. The total area in the CBS with DO concentration values of <3 mg/L was approximately 4.2 × 103 km2 (yellow dashed boxes in Fig. 1a). In the summer of the same year, Jiang et al. (2016) reported DO concentration values of <3 mg/L in the cold water mass near the southeastern BS, and the lowest value reached 2.3 mg/L (blue dashed boxes in Fig. 1a). They suggested that the long-term water stratification and the existence of more organic matter in the water has laid the foundation for the formation of the hypoxic zone. Using observational data obtained during the summer of 2019. Li et al. (2021) found that the area of hypoxia extended from the depression on the west side of the shoal in 2014 to the area around the mouths of the Huanghe River and the Laizhou Bay (green dashed boxes in Fig. 1a). The frequent occurrence and expansion of hypoxic events will directly affect the ecological environment in the CBS.
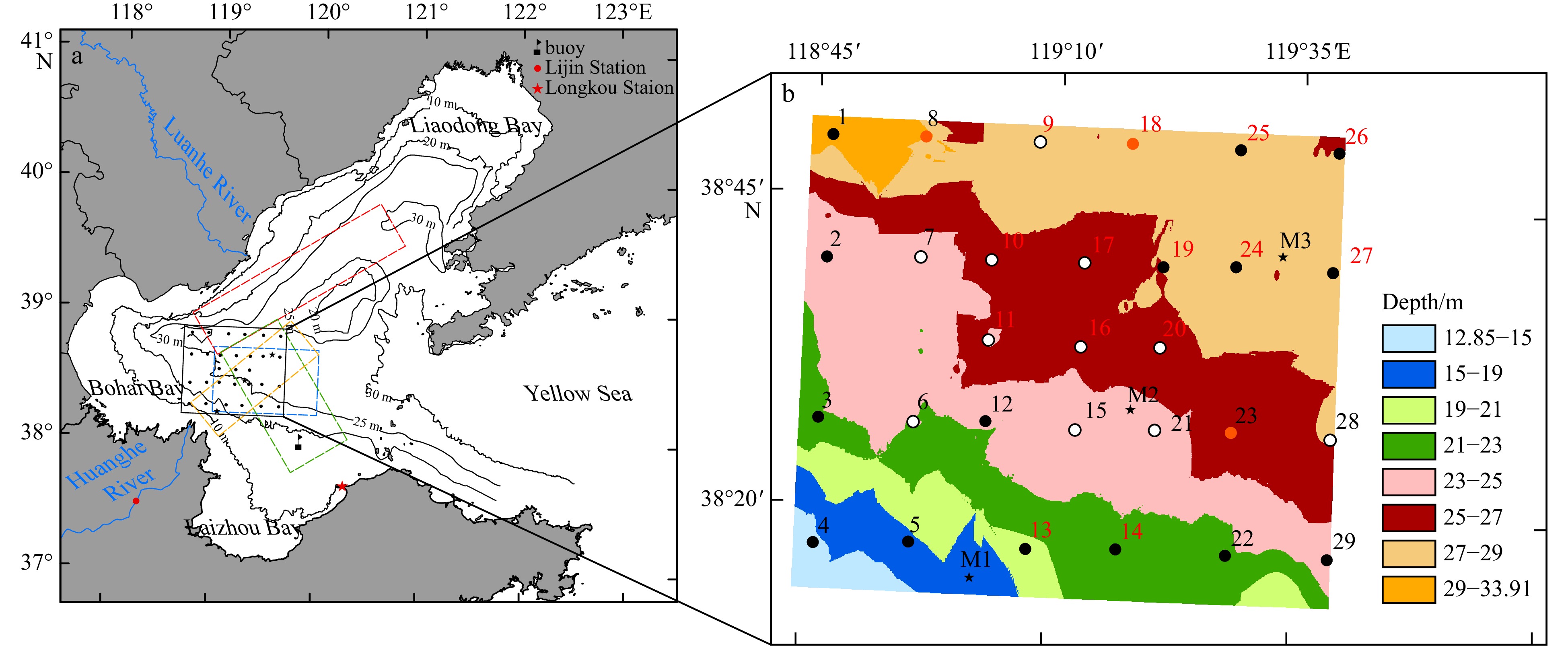
 Figure 1. Locations of survey stations in the CBS (Stas M1, M2, and M3). a. The black box is the study area; b. twenty-nine stations were distributed in the study area. In a, the yellow, blue, red, and green dashed boxes denote the hypoxic zones investigated by Zhang et al. (2016), Jiang et al. (2016), Zhai et al. (2012), and Li et al. (2021), respectively. The black contours and values indicate the water depth (in unit of m). In b, the black dots indicate DO concentration values of ≥4 mg/L, the white dots indicate DO concentration values of 3–4 mg/L, and the red dots indicate DO concentration values of <3 mg/L in summer in 2022; the black numbers indicate before strong winds, and the red numbers indicate after strong winds during the summer in 2022.
Figure 1. Locations of survey stations in the CBS (Stas M1, M2, and M3). a. The black box is the study area; b. twenty-nine stations were distributed in the study area. In a, the yellow, blue, red, and green dashed boxes denote the hypoxic zones investigated by Zhang et al. (2016), Jiang et al. (2016), Zhai et al. (2012), and Li et al. (2021), respectively. The black contours and values indicate the water depth (in unit of m). In b, the black dots indicate DO concentration values of ≥4 mg/L, the white dots indicate DO concentration values of 3–4 mg/L, and the red dots indicate DO concentration values of <3 mg/L in summer in 2022; the black numbers indicate before strong winds, and the red numbers indicate after strong winds during the summer in 2022.The hypoxic events in the CBS mentioned above all occurred in the area of the DCCWM. Zu et al. (2005) reported that the DCCWM was characterized by low temperatures and high salinity, and the DO concentration was lower than that in the surrounding area (Wei et al., 2019). Bao et al. (2004) suggested that the formation of DCCWM low-temperature zones may be closely related to the existence of a topographic trough in the BS and may result from the extension of the east and west low-temperature centers along this deep trough. Zhou et al. (2017) discovered the presence of a basin-scale cyclonic gyre induced by the DCCWM in summer in the BS, and the circulation potentially contributed to the development of hypoxia in this region.
The stratification of seawater is the hierarchical structure of the sea temperature (ST), salinity (SAL), and density (Den) of seawater with depth. Huang et al. (1999) suggested that seawater stratification is one of the causes of hypoxia in the BS. Stratification occurred in the BS from April to August, during which it gradually strengthened as the temperature increased and ended around mid-September (Huang et al., 1999; Zhou et al., 2017).
Based on discontinuous historical observational data from three stations in the CBS from 2013 to 2022, we studied the correlation between the stratification factor and the DO concentration via the linear regression method and analyzed the formation process of stratification during different months to determine the time at which the occurrence of hypoxia should be evaluated and verified. The results of this study enhance the understanding of the effect of stratification on hypoxia in the CBS and provide further references for early warning of hypoxia in the CBS.
2. Data and methods
2.1 Historical data
The historical data were obtained from the Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences (CAS), China, and were collected from Stas M1, M2, and M3 in the CBS (Fig. 1), which represent different water depths in the study area. The data included ST, SAL, Den, and DO concentration data, which were collected during a non-continuous comprehensive survey of the BS from 2013 to 2022. Observational data from the conductivity, temperature, and depth (CTD), SBE19+ CTD, and SBE25+ CTD (Sea-Bird Electronics, Inc., Bellevue, WA, USA) were used to determine the ST, SAL, and Den in situ. The DO concentration was measured using an SBE43 sensor. The field survey data were subjected to quality control and processing before being used in the subsequent analysis. The first step was to discard the ST, SAL, Den, and DO concentration data for outside the range boundaries provided in the user manual of the World Ocean Database 2018 [see Locarnini et al. (2018) Appendix 11 for min-max values of temperature, salinity, and DO concentration in coastal areas of the North Pacific]. The quality-controlled data were then interpolated to standard levels at 1 m vertical intervals. The air temperature (T) data were obtained from the Longkou Meteorological Station, and the Huanghe River runoff into the sea were obtained from the Lijin Hydrological Station. Sea surface temperature (SST) data from a buoy in the CBS were processed and analyzed by the Yantai Marine Center of the Ministry of Natural Resources, China.
2.2 Real-time survey data
The study area is located in the cold water mass in the southern BS (black rectangle in Fig. 1a). Real-time sea survey data (i.e., ST, SAL, Den, and DO concentration data) from May 28 to June 2 (June for short) and August 30 to September 4 (August for short), 2022, including data for the upper (0.5 m), middle (10 m), and bottom (2 m from the sea floor) three layers, from 29 stations, ranging from dense (Stas 10, 17, 19, 11, 16, 20, 12, 15, and 21) to scattered, were obtained. Stations intervals of 7.5 nautical miles horizontally and 6.5 nautical miles longitudinally in the dense area were established (Fig. 1b). The average distance between the other stations was approximately 10 nautical miles. The data were obtained using a YSI ProPlus multiparameter water quality analyzer (YSI, Inc., Yellow Springs, OH, USA) from late May through early June, as well as in August, in 2022. The SAL measurements were calibrated against standard seawater. The DO settings were calibrated using the water-saturated air method prior to each voyage.
2.3 Data from a base-type online monitoring system
A base-type online monitoring system (BTOMS) was deployed at Sta. M2 (Fig. 1b) on the seabed in the CBS from June to August in 2023. The data were acquired (Table 1) at an interval of 1 h. The BTOMS was equipped with a YSI EXO2 multiparameter water quality analyzer, which contained a depth sensor (strain gauge pressure principle), DO sensor (fluorescence method), total algae sensor (fluorescence method), conductivity/temperature composite sensor (thermistor principle), and pH sensor (glass composite electrode method).
Table 1. Data sources of historical and real-timeData classification Time Station Data Historical data 2013–2022 M1, M2, M3 ST, SAL, Den, DO concentration 2011–2022 Longkou Station T 2010–2022 Lijin Station Huanghe River runoff Real-time data June and August in 2022 Stas 1–29 ST, SAL, Den, DO concentration June–August in 2023 M2 ST, SAL, Den, DO concentration June–August in 2023 buoy SST, SAL | Show Table DownLoad:
CSV
DownLoad:
CSV
2.4 Stratification factors and DO characteristic analysis methods
The regression analysis method (RAM) was used to analyze the historical data. Taking data from Sta. M2 as an example, the characteristics of the stratification factors (ST, SAL, and Den) and DO concentration over time were analyzed. The month of the initial formation of the stratification was also determined. A general global optimization algorithm was used to simulate the correlations between the DO concentration and the ST, SAL, and Den with depth and to simulate the correlations between the DO concentration and the ST, SAL, and Den values of the upper layer minus the values of the bottom layer (Δ) from 2013 to 2022. The roles of the stratification factors in the hypoxic events was also determined. The thresholds of the stratification factors during the month in the early formation stage of the stratification during hypoxic events were determined. The method of determining the thresholds of the stratification factors for Stas M1 and M3 was the same as that for Sta. M2. The thresholds of the stratification factors at these three stations were applied to the survey data for 2022 and 2023 to estimate the likelihood of the occurrence of hypoxia at each station. The workflow of our research is shown in Fig. 2c.
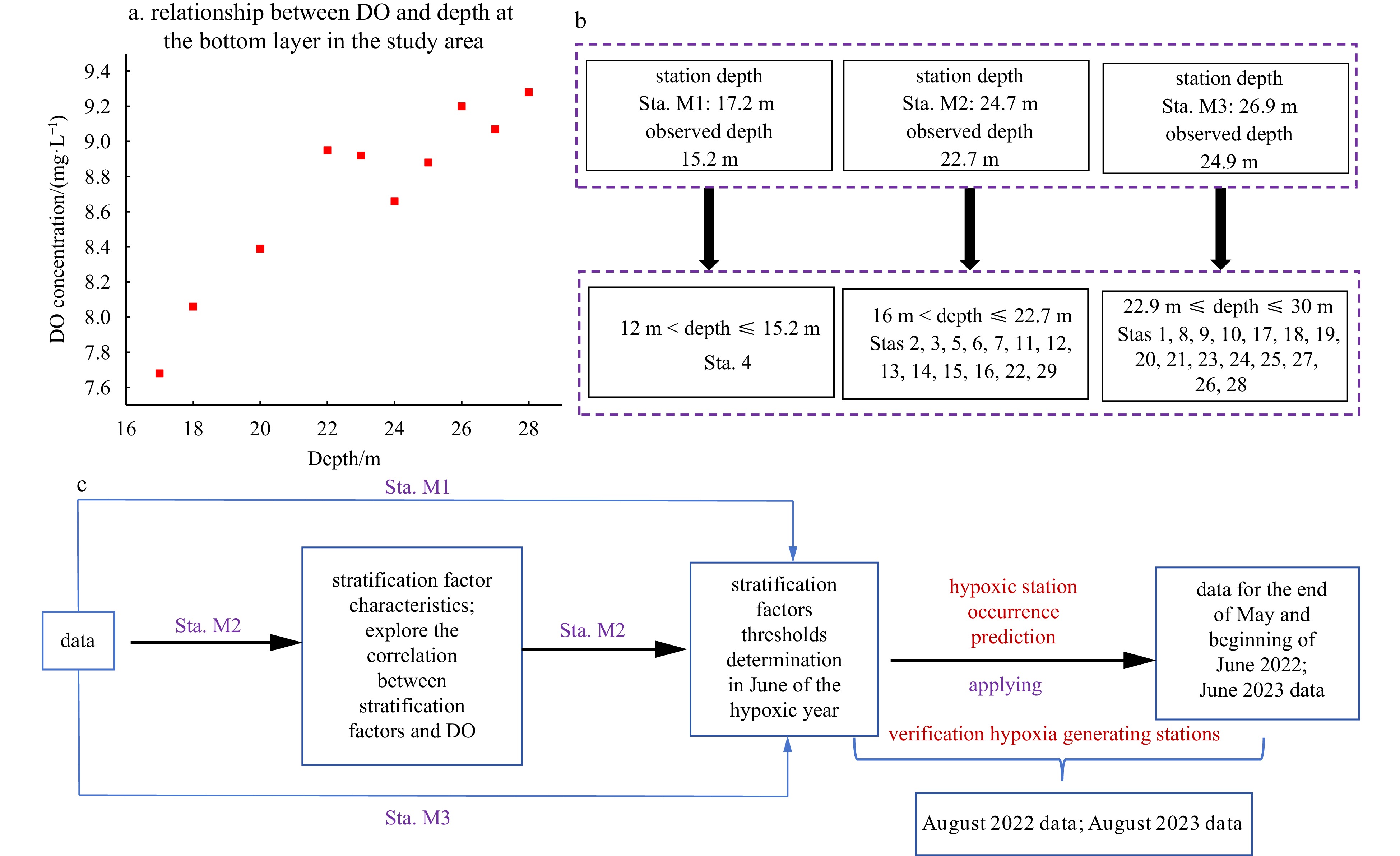
 Figure 2. Research methods: a. the correlation between the bottom DO concentration and water depth in the study area in spring 2022; b. the observation depth range for the 29 stations (Stas 1−29) in the study area corresponds to that of Stas M1, M2, and M3; and c. the workflow of our research.
Figure 2. Research methods: a. the correlation between the bottom DO concentration and water depth in the study area in spring 2022; b. the observation depth range for the 29 stations (Stas 1−29) in the study area corresponds to that of Stas M1, M2, and M3; and c. the workflow of our research.2.5 Observation depths of the 29 stations matched with Stas M1, M2, and M3
As shown in Fig. 2a, there was a correlation between the DO concentration values at the bottom of the study area and the depth at the same station in spring in 2022. The different depths of the historical data for Stas M1, M2, and M3 were collected in the study area and the sites gradually deepened from southwest to northeast (Fig. 1b). Because the depth of the observations was usually approximately 2 m from the seabed, in order to better compare the analysis data with the historical data, we matched the observed depths of the 29 stations with the observed depths close to Stsa M1, M2, and M3 (Fig. 2b).
2.6 Hypoxic environment threshold settings in the study area
The seawater quality was divided into Class 1 (DO concentration > 6 mg/L), Class 2 (DO concentration > 5 mg/L), Class 3 (DO concentration > 4 mg/L), and Class 4 (DO concentration > 3 mg/L). Classes 1 and 2 were good water bodies, that is, healthy water bodies. In contrast, water bodies with DO concentration values of <4 mg/L were unhealthy water bodies (National Standard of the People’s Republic of China: Standard for Seawater Quality, UCD 551463 GB3097-1997). In this study, environments with DO concentration value of <4 mg/L were defined as a hypoxic environments in order to predicate hypoxic events.
Some datasets for this research are included in this paper and will be made available upon request at
https://www.scidb.cn/ .3. Results
3.1 Distribution characteristics of stratification factors and DO concentration at Sta. M2 in the CBS
The data were derived from the accumulated voyage survey data at Sta. M2 from 2013 to 2022 (discontinuous, no fixed survey period) (Fig. 3). To achieve a better comparison, the data for December, February (dotted line), and April–June (*line) are plotted in Figs 3a, c, e, and g, and the data for August (line) and November (○line) are plotted in Figs 3b, d, f, and h. Figures 3a and b show that the ST from the upper and bottom layers in the CBS were basically uniform in December and February. The ST stratification began in April, and an obvious thermocline formed in June. In August, the thermocline strengthened further and the water depth was 5–10 m, the maximum ST difference in the thermocline was 5.5℃, and the ST outside the thermocline only changed slightly. The stratification gradually disappeared after August.
 Figure 3. Variations in the ST, SAL, Den, and DO concentration with depth in different seasons at Sta. M2 in the CBS from 2013 to 2022. a. ST from winter and spring, b. ST from summer and autumn, c. SAL from winter and spring, d. SAL from summer and autumn, e. density from winter and spring, f. density from summer and autumn, g. DO concentration from winter and spring, and h. DO concentration from summer and autumn. To achieve a better comparison, the data for December, February (dotted line), and April–June (*line) are plotted in Figs 3a, c, e, and g, and the data for August (line) and November (○line) are plotted in Figs 3b, d, f, and h.
Figure 3. Variations in the ST, SAL, Den, and DO concentration with depth in different seasons at Sta. M2 in the CBS from 2013 to 2022. a. ST from winter and spring, b. ST from summer and autumn, c. SAL from winter and spring, d. SAL from summer and autumn, e. density from winter and spring, f. density from summer and autumn, g. DO concentration from winter and spring, and h. DO concentration from summer and autumn. To achieve a better comparison, the data for December, February (dotted line), and April–June (*line) are plotted in Figs 3a, c, e, and g, and the data for August (line) and November (○line) are plotted in Figs 3b, d, f, and h.The SAL distribution is shown in Figs 3c and d. In December and February, the SAL of the subsurface layer increased, after which the SAL changed little with depth; SAL stratification initially appeared in April. The SAL was strongly stratified due to external influences in April 2022. The variation trend of halocline in June, August, and after August were the same as that of the thermocline. The halocline was mainly stable in water depths of 5–10 m, the maximum SAL difference was 0.6, and the SAL only changed slightly in the regions outside the halocline.
Both the thermocline and halocline gradually strengthened with increasing sea surface temperature. Because the density of sea water is related to the SST and salinity, the pycnocline, thermocline, and halocline had the same change trends in April, June, August, and after August. The pycnocline formed at a depth of 5–10 m (Figs 3e and f).
Figures 3g and h show that the change in the loss of DO concentration with depth was not obvious in December, February, April, and May. It began to become obvious in June, and the trend of the loss of DO concentration further increased in August. At depths of 5–10 m, the DO concentration value exhibited a decreasing trend; and at depths of 10–15 m, the DO concentration value exhibited a larger decreasing trend, that is, beneath the stratification of seawater (at a depth of 5–10 m below), the DO concentration value exhibited a large trend of depletion at depths of 10–15 m.
It was found that seawater stratification initially formed in June, and the oxygen consumption at the bottom initially appeared in June. In August, the stratification was enhanced and the oxygen consumption in the bottom layer was enhanced.
3.2 Correlation analysis between DO concentration and ST, SAL, and Den at Sta. M2 in the CBS
A total of 79 sets of data were obtained for February, June, August, and December in 2017. The simulation results were evaluated using the correlation coefficient (R), root mean square error (RMSE), residual sum of squares (SSE), R-squared, and the determination coefficient (DC) using a general global optimization algorithm.
The correlations between the simulated stratification factors and DO concentration with seawater depth (depths of 1–25 m, n = 79) are shown in Table 2. The correlation coefficients of the ST, SAL, and Den with DO concentration are 0.91, 0.45, and 0.81, respectively. The RMSE values are 0.97 mg/L, 2.06 mg/L, and 1.34 mg/L, respectively. The R-squared values are 0.84, 0.20, and 0.66, respectively. The simulation of the correlation between the ST and DO concentration are the best, followed by the results of the correlation between the DO concentration and the Den, and the results for the correlation between the DO concentration and the SAL are the worst.
Table 2. Simulation results of the correlations between the stratification factors and DO concentrationCorrelation ST/℃ and DO
concentration/(mg·L–1)SAL and DO
concentration/(mg·L–1)Density/(kg·m–3) and
DO concentration/(mg·L–1)y DO concentration DO concentration DO concentration x ST SAL Density Formula Y = 10.482 − 0.002x2.5 + 4.42 × exp(x) Y = 10.08 − 3.6/{1 + [(x − 31.54)/0.14]2} y = 5.7 × 106 − 1.67 × 104x + 16.27 × x2 − 5.3 × 10−3x3 R 0.91 0.45 0.81 RMSE/
(mg·L–1)0.97 2.06 1.34 SSE 74.62 335.06 141.52 R-squared 0.84 0.20 0.66 DC 0.82 0.20 0.66 Correlation ΔST/℃ and ΔDO
concentration/(mg·L–1)ΔSAL and ΔDO
concentration/(mg·L–1)ΔDen/(kg·m–3) and ΔDO
concentration/(mg·L–1)y ΔDO concentration ΔDO concentration ΔDO concentration x ΔST ΔSAL ΔDen Formula y = 3.36 × exp[−exp(110.06 – 47.53 x)] y = exp[−25.74 + 82.61x3+21.82exp(−x)] y = (0.69 + x)/(8.04 + 7.84x) − 1.05x R 0.82 0.78 0.81 RMSE/
(mg·L–1)0.96 1.05 0.97 SSE 17.64 20.77 17.99 R-squared 0.66 0.61 0.67 DC 0.66 0.60 0.66 | Show TableDownLoad:
CSV
The simulated correlations between the ΔDO and the ΔST, ΔSAL, and ΔDen are shown in Table 2. The R values of the correlations between the ΔDO and the ΔST, ΔSAL, and ΔDen are 0.82, 0.78, and 0.81, respectively, which are close to each other. The RMSE values are 0.96 mg/L, 1.05 mg/L, and 0.97 mg/L, respectively, and the R-Squares values are 0.66, 0.61, and 0.67, which are relatively close.
Based on a comparison of the simulation results, it is reasonable to use the ΔST, ΔSAL, and ΔDen values to evaluate the occurrence of hypoxia. Stratification plays a leading role in the occurrence of hypoxia at Sta. M2.
3.3 Thresholds of stratification factors at Sta. M2
In the CBS, stratification occurred in June and was further strengthened in August, in which the largest DO concentration loss occurred. In this section we determine the stratification factor thresholds in June during hypoxia years, and we pre-analyze the possibility of hypoxic events occurring in August.
We analyzed historical data and published articles for the years in which hypoxia occurred in summer (2014, 2015, 2017, 2019, and 2020) in the CBS. As can be seen from Fig. 3h, the minimum DO concentration value (2.37 mg/L) of the bottom layer in the CBS occurred in August 2017.
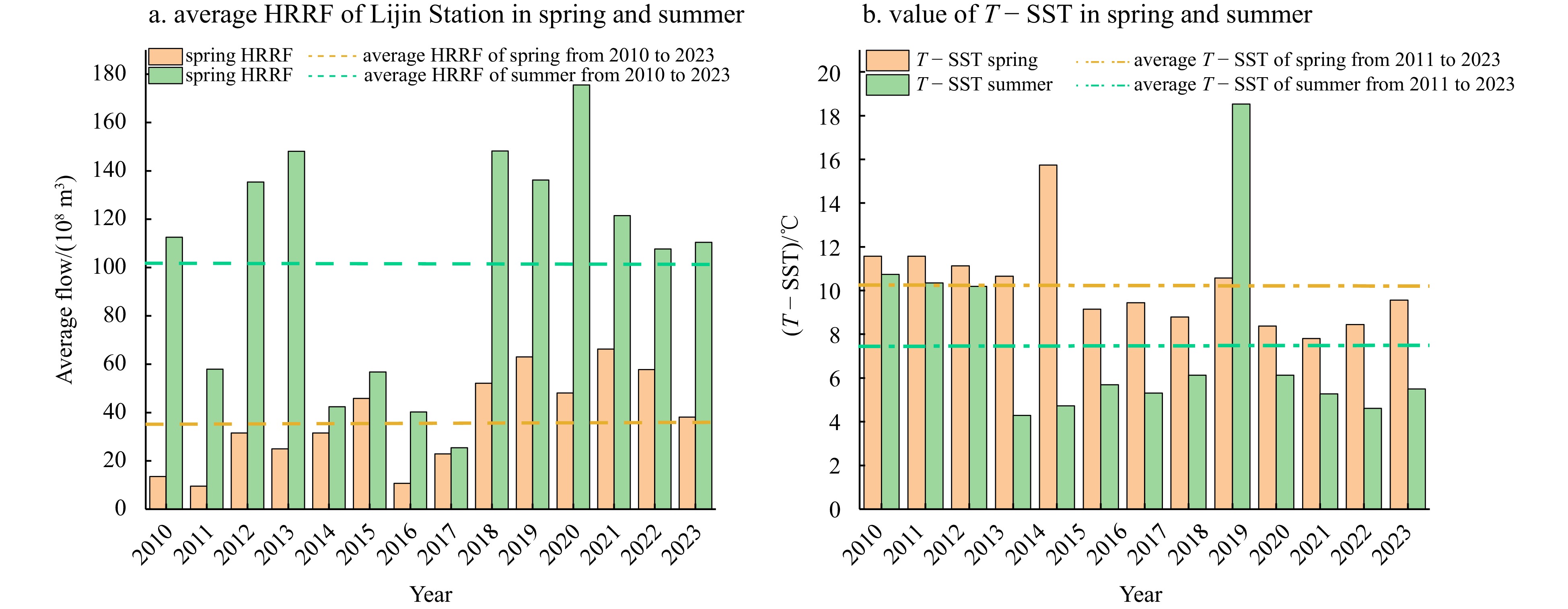
Due to the discontinuous nature of the observational data, we only had stratification factor data for hypoxic events in June in 2017 and 2019 (Fig. 3). In June 2017, the values of the stratification factors (ΔST, ΔSAL, ΔDen, and ΔDO) were 2.32℃, −0.24, −0.83 kg/m3, and 1.45 mg/L, respectively. In June 2019, the values of these stratification factors were 4.04℃, −0.49, −1.37 kg/m3, and 0.41 mg/L, respectively. The absolute values of the stratification factors were all higher in 2019 than in 2017, but the ΔDO value was much smaller in 2019 than in 2017. Figure 4a shows the observed Huanghe River runoff in spring and summer at Lijin Station in the Huanghe River Estuary from 2010 to 2023. In both the spring and summer in 2017, the Huanghe River runoff (HRRF) was lower than the multi-year seasonal average, indicating that the hypoxia in summer in 2017 was less affected by the HRRF into the sea. Figure 4b shows the difference between the air temperature T at Longkou Station and the SST at the buoy in the CBS during spring and summer from 2011 to 2023. The difference between T and SST in the spring and summer in 2017 was lower than the multi-year seasonal average and was the smallest, indicating that T and SST in 2017 were less affected by the unusual weather. The values in 2019 were greater than the multi-year average. Therefore, we selected the stratification factors in June 2017 as thresholds (ΔST ≥ 2.32℃, ΔSAL ≤ −0.24, and ΔDen ≤ −0.83 kg/m3) to assess that the hypoxia in August in the CBS was reasonable.
 Figure 4. HRRF at Lijin Station and the air temperature T at Longkou Station minus the SST at the buoy statistics: a. the HRRF of spring and summer at the Lijin Station from 2010 to 2023, and b. the difference between T at Longkou Station and the SST at the buoy in the CBS in spring and summer from 2011 to 2022.
Figure 4. HRRF at Lijin Station and the air temperature T at Longkou Station minus the SST at the buoy statistics: a. the HRRF of spring and summer at the Lijin Station from 2010 to 2023, and b. the difference between T at Longkou Station and the SST at the buoy in the CBS in spring and summer from 2011 to 2022.3.4 Characteristics and thresholds of the stratification factors at Stas M1 and M3
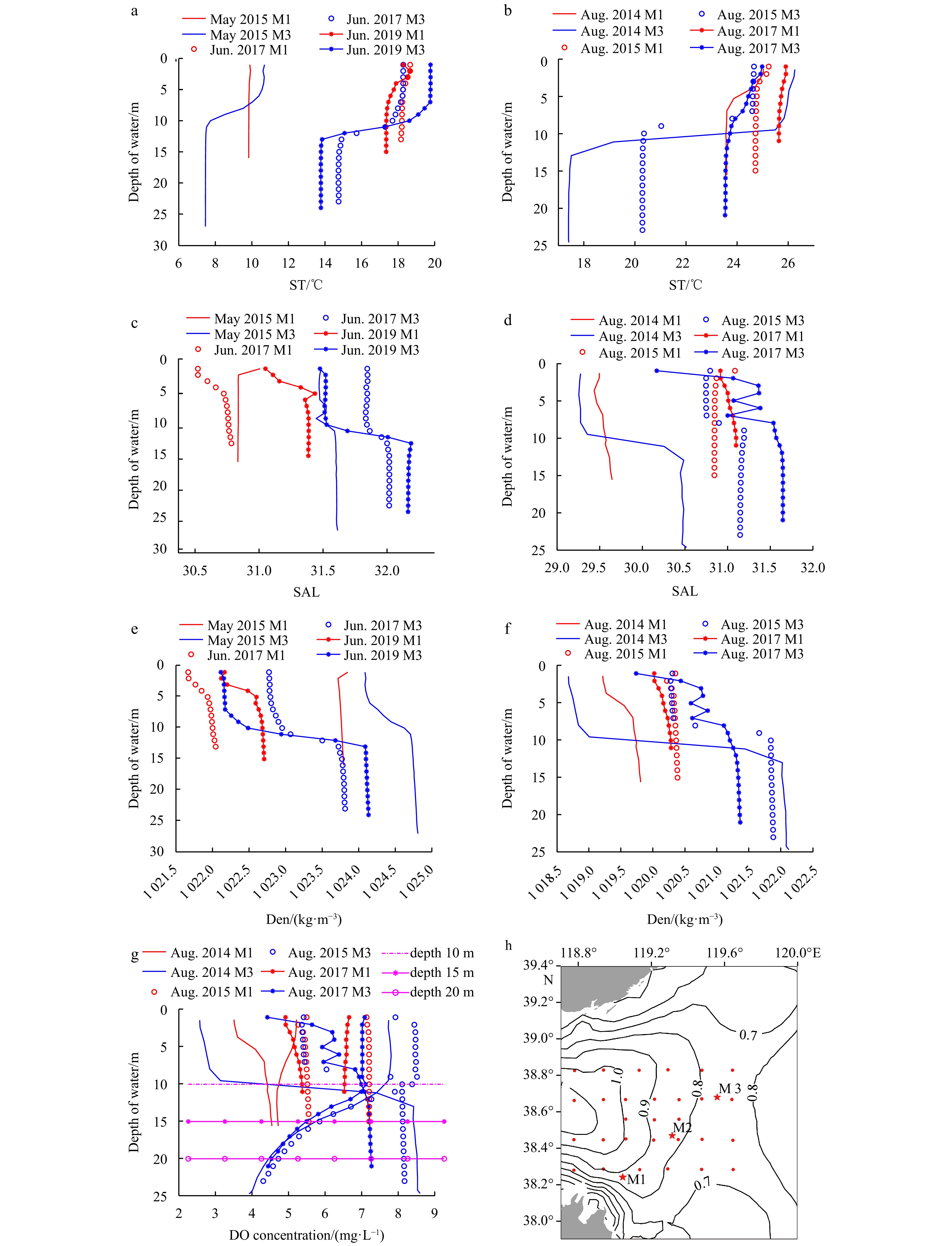
In Figs 5a and b, the red line represents the ST value at Sta. M1. As the water depth at Sta. M1 (17.2 m) is shallow, the ST stratification was not obvious in June or August. The thermocline at Sta. M3 initially formed in June and was further strengthened in August. The thermocline was mainly llocated at depths of 10–15 m. Figures 5c and e show that the SAL and Den at Sta. M1 were stratified in June, and the depths of the stratifications were all 0–5 m. The SAL stratification disappeared in August (Fig. 5d), and the Den stratification did not strengthen (Fig. 5f). As a result, in August, the DO concentration (Fig. 5g) at Sta. M1 did not reach hypoxic conditions (DO concentration < 4 mg/L). The development of the halocline (Figs 5c and d) and density cline (Figs 5e and f) at Sta. M3 in June and August was consistent with that of the thermocline. As can be seen from Fig. 5g, at Sta. M3, the DO concentration decreased at depths of 10–15 m in the stratified interval, and it decreased faster below the stratified interval. Comparison of the data for Stas M2 and M3 revealed that they were both located in areas with low flow velocities (Fig. 5h). The current velocity around Sta. M1 was between 0.7 m/s and 0.9 m/s (Zheng et al., 2021). The depth was 2.2 m deeper at Sta. M3 than at Sta. M2, resulting in the depth of the stratification layer at Sta. M3 being lower than that at Sta. M2. The determination of the stratification factor threshold for Sta. M3 in June was also based on the data for 2017, and the method of determining the threshold value was the same as that for Sta. M2. The stratification factor thresholds (SFTs) for the three stations at different depths in the study area are shown in Table 3.
 Figure 5. Information about the stratification factors and DO concentration at Stas M1 and M3 and distribution of flow field in the CBS. a. ST distribution with depth in May and June in 2015, 2017, and 2019; b. ST distribution with depth in August in 2014, 2015, and 2017; c. SAL distribution with depth in May and June in 2015, 2017, and 2019; d. SAL distribution with depth in August in 2014, 2015, and 2017; e. Den distribution with depth in May and June in 2015, 2017, and 2019; f. Den distribution with depth in August in 2014, 2015, and 2017; g. DO concentration distribution with depth in August in 2014, 2015, and 2017; and h. the maximum tidal current velocity in May 2022. In h, red dots: stations; red star: Stas M1, M2 and M3; contour lines and numbers represent the velocity (m/s) of the flow.Table 3. SFTs for Stas M1, M2, and M3 in June during a hypoxia year
Figure 5. Information about the stratification factors and DO concentration at Stas M1 and M3 and distribution of flow field in the CBS. a. ST distribution with depth in May and June in 2015, 2017, and 2019; b. ST distribution with depth in August in 2014, 2015, and 2017; c. SAL distribution with depth in May and June in 2015, 2017, and 2019; d. SAL distribution with depth in August in 2014, 2015, and 2017; e. Den distribution with depth in May and June in 2015, 2017, and 2019; f. Den distribution with depth in August in 2014, 2015, and 2017; g. DO concentration distribution with depth in August in 2014, 2015, and 2017; and h. the maximum tidal current velocity in May 2022. In h, red dots: stations; red star: Stas M1, M2 and M3; contour lines and numbers represent the velocity (m/s) of the flow.Table 3. SFTs for Stas M1, M2, and M3 in June during a hypoxia yearItem Value Sta. M1 Sta. M2 Sta. M3 ΔST/°C (assessment SFTs of hypoxia in June ) − ≥2.32 ≥3.54 ΔSAL (assessment SFTs of hypoxia in June) − ≤−0.24 ≤−0.17 ΔDen/(kg·m–3) (assessment SFTs of hypoxia in June) − ≤−0.83 ≤−1.04 Note:− indicates that there is no threshold. | Show TableDownLoad:
CSV
3.5 Application of SFTs
A survey was conducted in the CBS from June, 2022, at 29 stations in the upper, middle, and lower layers (Fig. 1b). ST, SAL, Den, and DO concentration data for 29 stations were obtained. The SFTs in June (Table 3) were applied to the stratification factors at 29 stations in June 2022. According to the results presented in Fig. 2b, the observation depths at the 29 stations were matched with the data for Stas M1, M2, and M3, and the SFTs determined using these three stations in June were used to predict whether hypoxia occurred in August at the 29 stations. The evaluation results are shown in Table 4. Only Sta. 4 corresponded to the depth at Sta. M1, and the possibility of hypoxia occurring was very small and thus is not listed on the Table 4. Thirteen stations were judged based on the Sta. M2 SFTs, and seven stations (Stas 7, 11, 12, 14, 15, 16, and 22) reached these SFTs in June. Meanwhile, 15 stations were evaluated based on the Sta. M3 SFTs, and 12 stations (Stas 9, 10, 17, 18, 19, 20, 21, 23, 24, 25, 27, and 28) reached these SFTs in June. That is, according to the stratification factor assessment in June 2022, 19 stations would experience hypoxia in August.
Table 4. Assessment of the stratification factors for June, 2022Station Sta. M2 Station Sta. M3 ΔST ≥ 2.32 ℃ ΔSAL ≤ −0.24 ΔDen ≤ −0.83 kg/m3 ΔST > 3.5 ℃ SAL ≤ −0.17 ΔDen ≤ −1.04 kg/m3 2, 3 √ × √ 1 √ × √ 5 × × × 8, 26 × × × 6, 29 √ × √ 9, 10, 17 √ √ √ 7, 11, 12 √ √ √ 18, 19, 20 √ √ √ 13 √ × × 21, 23, 24, √ √ √ 14, 15 √ √ √ 25, 27, 28 √ √ √ 16, 22 √ √ √ 29 √ × × Note: √ indicates that the stratification factor thresholds are satisfied. × indicates that the stratification factor thresholds are not satisfied. | Show TableDownLoad:
CSV
A summer survey was conducted from August 30 to September 4, 2022, in the CBS. However, between August 31 and September 2, gale force winds of 6 (Beaufort scale: wind speed between 10.8 m/s and 13.8 m/s) and above occurred, and the ship was sheltered in the port. The DO concentration and ▲DO values (the bottom layer DO concentration in spring minus the bottom layer DO concentration in summer) of the bottom layer at the 29 stations are shown in Fig. 6. A total of 14 stations (Stas 6, 7, 8, 9, 10, 11, 15, 16, 17, 18, 20, 21, 23, and 28) had DO values of <4 mg/L of which three stations (Stas 8, 18, and 23) had DO concentration values of <3 mg/L, and the minimum DO concentration value occurred at Sta. 8 (DO = 2.92 mg/L) (Fig. 6a). Twelve stations confirmed that the June assessment had an accuracy of 63%. As can be seen from Fig. 6b, the ▲DO values at the hypoxic stations were basically greater than 4 mg/L (except for Sta. 28, ▲DO = 3.92 mg/L). The hypoxic zone was boot-shaped.
 Figure 6. Hypoxic validation of the August 2022 and 2023 data in the CBS. a. DO concentration distribution in the bottom layers at the 29 stations in August 2022 (black numbers: DO concentration values); b. distribution of bottom DO concentration values in June 2022 minus bottom DO concentration values (▲DO) in August at the 29 stations (black numbers:▲DO concentration values); c. comparison of ▲stratification factors (surface value minus bottom value) and SFTs (dotted line) in June 2023; and d. DO concentration distribution (the dash line is DO concentration = 4 mg/L) at the bottom at Sta. M2 during August 1–20, 2023.
Figure 6. Hypoxic validation of the August 2022 and 2023 data in the CBS. a. DO concentration distribution in the bottom layers at the 29 stations in August 2022 (black numbers: DO concentration values); b. distribution of bottom DO concentration values in June 2022 minus bottom DO concentration values (▲DO) in August at the 29 stations (black numbers:▲DO concentration values); c. comparison of ▲stratification factors (surface value minus bottom value) and SFTs (dotted line) in June 2023; and d. DO concentration distribution (the dash line is DO concentration = 4 mg/L) at the bottom at Sta. M2 during August 1–20, 2023.A BTOMS was deployed on the seabed at Sta. M2 (Fig. 1b) in the CBS from June to August 2023. Although the ST, SAL, and Den of sea surface were not measured by the BTOMS at Sta. M2, the observed values at the buoy near the study area are representative to a certain extent. Figure 6c shows the stratification factors (ΔST, ΔSAL, and ΔDen) of the sea surface values observed by the buoy minus the seafloor values observed by the BTOMS in June 2023. The dotted line represents the SFTs at Sta. M2 in June (Fig. 6c). The stratification factors from the buoy and BTOMS fully satisfy the SFTs. It was predicted that a hypoxic event would occur at Sta. M2 in August. Figure 6d shows the valid DO concentration data for August 1–20, 2023, some of the DO concentration values were <4 mg/L from August 2 to 12. The DO concentration gradually decreased from August 12 to 20, and all of the values were lower than 4 mg/L. Hypoxic events occurred at Sta. M2 in August 2023.
4. Discussion
4.1 Validity and rationality of the historical data for the CBS
Despite the discontinuous nature of the observed data in the CBS and the limited range of the data, it was determined that the stratification in the CBS basically formed in June, and the intensification in August was the best time to observe the hypoxia. Stratification occurred in the CBS from April to August. During this time, it gradually strengthened as the temperature increased. This is consistent with the results of Huang et al. (1999) and Zhou et al. (2017).
The correlation analysis of the stratification factors and DO concentration with depth revealed that the correlation coefficient between the SAL and DO concentration was only 0.45, the RMSE was 2.06 mg/L, and the R-squared value was 0.20, which were caused by the flow and sediment regulation of the nearshore rivers, especially the Huanghe River. However, the correlation coefficients of the ΔDO and the ΔST, ΔSAL, and ΔDen were close to R ≥ 0.78, RMSE ≤ 1.05, and R-squared ≥ 0.61. Therefore, it is reasonable to evaluate the occurrence of hypoxic events using the differences in the stratification factors (ΔST, ΔSAL, and ΔDen) between the surface layer and the bottom layer.
In addition, consistent with the historical classification of the water health status in the BS, the data analysis, and published results, the stratification factors (ΔST, ΔSAL, and ΔDen) were found to be the principal factors assessing hypoxia in the CBS, and the hypoxic environment was defined as DO concentration < 4 mg/L, which is in line with the characteristics of the BS. As the spawning and breeding periods of economically important fish, shrimp, and crab in the BS are between March and August. The water dynamics are weak in summer, and the increases in the sea temperature and stratification prevent exchange between the water in the upper and lower layers. Frequent hypoxic events in the CBS threaten fishery resources in the Yellow and Bohai seas.
4.2 Analysis of the factors affecting the evaluation of hypoxia in the CBS in 2022
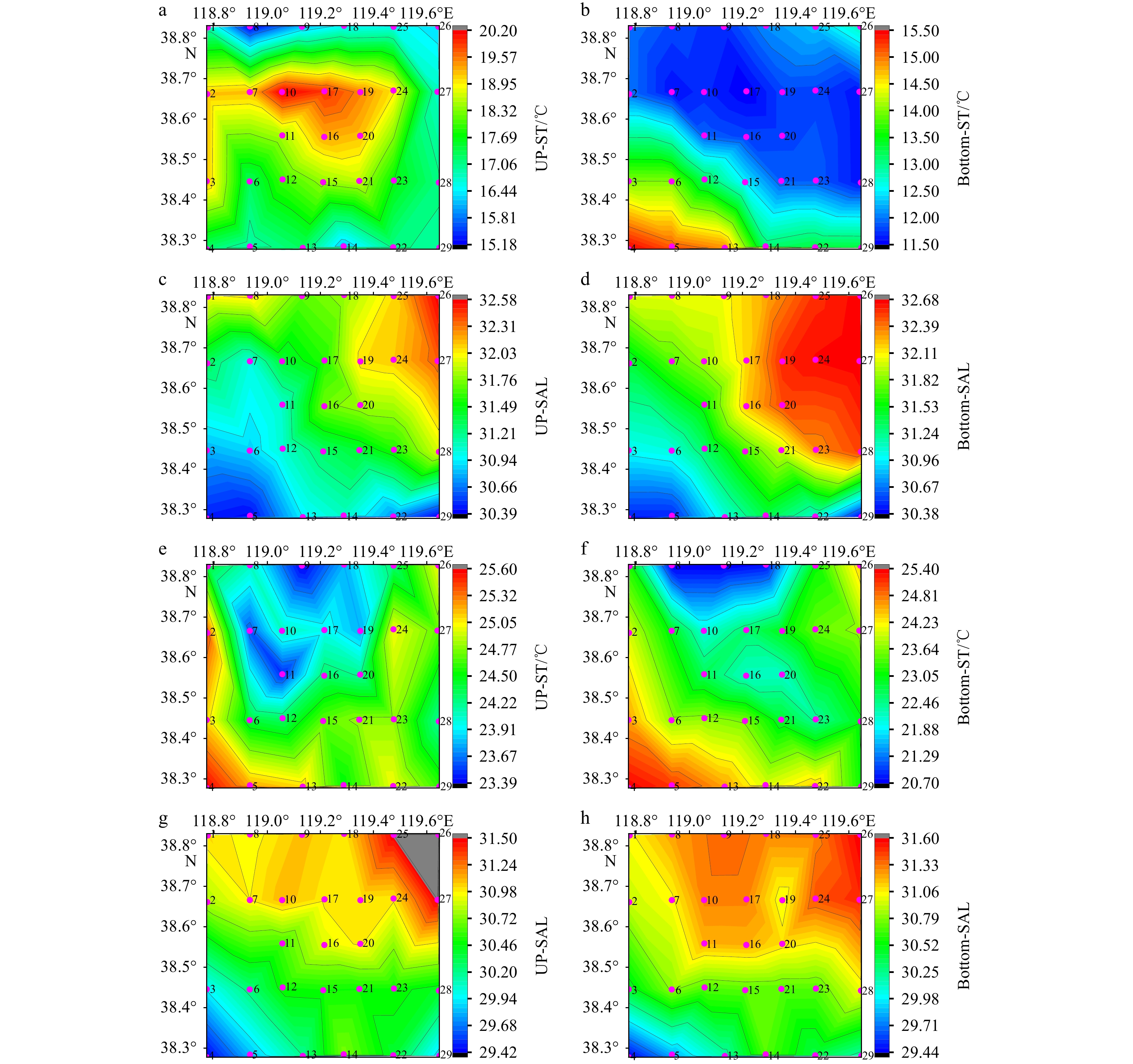
According to the evaluation and verification of the hypoxia in Section 3.5, hypoxia will occur at 19 stations in the study area. As a result, hypoxia occurred at 14 stations, of which 12 from 19 stations. Stations 6 and 8 were hypoxic stations in August, but they were not in evaluation stations. In particular, the ΔST, ΔSAL, and ΔDen at Sta. 8 did not meet the SFTs in June. However, in August, Sta. 8 was the hypoxic station with the lowest DO concentration value in the study area. As can be seen from Figs 7a and c, the surface layer at Sta. 8 received low ST and high SAL water from the north during the survey in early June, which interfered with the prediction. For the survey conducted at the end of August, it can be seen from Figs 7f and h that low ST and high SAL water entered the bottom layer near Stas 8, 9, and 18 from the north, resulting in enhanced stratification, which led to hypoxia at Sta. 8. As the water depth at Sta. 8 was the greatest among the stations with hypoxia, the DO concentration value of its bottom layer was the smallest during August. Due to the time lag of the formation of stratification at Sta. 8, the June forecast failed. As can be seen from Figs 7a–d, Sta. 6 was greatly affected by near-shore influences. In particular the SAL was influenced by the nearshore rivers, resulting in an excessive ΔSAL value in June, which could not be accurately evaluated.
 Figure 7. ST and SAL distributions in June and August in 2022: a. up-ST in June; b. bottom-ST in June; c. up-SAL in June; d. bottom-SAL in June; e. up-ST in August; f. bottom-ST in August; g. up-SAL in August; and h. bottom-SAL in August. The pink dots and numbers represent the stations.
Figure 7. ST and SAL distributions in June and August in 2022: a. up-ST in June; b. bottom-ST in June; c. up-SAL in June; d. bottom-SAL in June; e. up-ST in August; f. bottom-ST in August; g. up-SAL in August; and h. bottom-SAL in August. The pink dots and numbers represent the stations.A summer survey was conducted from August 30 to September 4, 2022, in the CBS. However, between August 31 and September 2, gale force winds of 6 (Beaufort scale) and above occurred, and the ship was sheltered in the port. In Fig. 1b, the black station numbers indicate the period before the strong winds, and the red station numbers indicate the period after the strong winds during the summer of 2022. Stations 14, 24, 25, and 27 were investigated after the occurrence of strong winds. Although they were the stations that were about to experience hypoxia in the assessment in June, the survey data for August revealed that the DO concentration values were >4 mg/L and hypoxia did not occur, which should have been caused by the strong winds stirring the water and causing the DO concentration to rise. As can be seen from Fig. 7, the southwestern part of the study area was greatly affected by the nearshore area, the northeastern part was affected by the central bank of the BS, the western part was affected by Bohai Bay, the southern part was affected by Laizhou Bay, and the northwestern part was affected by the entry of low ST and high SAL water. In accordance with these observations, it was estimated in June that hypoxia was about to occur at Stas 12, 19, and 22; however, the data from August indicated that hypoxia did not occur at these stations.
5. Conclusions
In this study, we analyzed historical data for Stas M1, M2, and M3 in the CBS. The stratification of the CBS started in April, and it initially formed in June, which can be used to evaluate the occurrence of future hypoxic events. The stratification was strongest in August, which is the best time to observe the occurrence of hypoxia. It was found that it is feasible to predict the occurrence of hypoxia in August using the ΔST, ΔSAL, and ΔDen thresholds from June with the RAM analysis method. Thus the hypoxia SFTs in June were determined from the data Stas M1, M2, and M3, and the possibility of hypoxia occurring in the CBS in August 2022 and August 2023 was evaluated and verified using the measured data. It was found that stratification played a leading role in the occurrence of hypoxic events in the CBS. The stratification in the CBS was related to the seabed topography. The inflow of coastal rivers into the sea, windy days when hypoxia occurred, Bohai Bay, Laizhou Bay, the central bank of the BS, and the inflow of low temperature and high salinity water affected the stratification factors in the study area and interfered with the accuracy of the assessment.
Based on the analysis and assessment using data from 29 stations in the CBS at the end of May to early June in 2022, hypoxia will occur at 19 stations. The verification of the survey data from 29 stations at the end of August revealed the occurrence of hypoxia at 14 stations, of which 12 were evaluation stations, and the overall accuracy was 63%. Severe oxygen depletion occurred in the deep water area during August in 2022 in the CBS, the hypoxic values varied from 2.92 mg/L to 3.93 mg/L, and the depths of the hypoxic stations were greater than 22 m. The observation data for the area around Sta. M2 collected by a BTOMS in June 2023 were used to evaluate the possibility of hypoxia occurring in August based on the SFTs in June, and the observation data for August of the same year were used to verify that hypoxic events occurred in this region.
This method can only be used to evaluate hypoxic events with DO concentration values of <4 mg/L and cannot be used to characterize the severity of the hypoxia. Biological mineralization and degradation can promote the development of hypoxia. In the future, perhaps consideration of stratification, biodegradation, and mineralization can be used solve the severity of hypoxia.
Acknowledegements
This work was supported by the Data Center of Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, China. Some of the data and samples were collected utilizing R/V Lanhai101 during open research cruise NORC2023-01, supported by the NSFC Shiptime Sharing Project under contrac No. 42249901.
-

Figure 1. Light microscopy (LM) and scanning electron microscopy (SEM) images of P. clipeus DF128. Light microscopy, right thecal view showing the cell shape, the large nucleus (N) posterior (a). Laser scanning confocal microscopy images of P. clipeus (b–d). Epifluorescence image showing the pyrenoid (Py) and radial arrangement of chloroplasts (Chl) (b). Sybr Green stained cell showing the shape of the nucleus (N) (c). Epifluorescence image of the thecal valve of the cell (d). The right valve view shows that the cell shape is asymmetrical and round (SEM) (e). Left valve view showing the smooth thecal surface with a radial pore pattern (SEM) (f). The intercalary band is wide and has transverse striation (SEM) (g). The cell is shown in the right lateral view, with a wide arc-shaped periflagellar area. Ridge (asterisk), wing-shaped protrusion (arrow), curved projections (arrowhead), detail of the nine platelets (SEM) (h and i). Scale bars in a–g: 10 μm, h and i: 5 μm.

Figure 2. Maximum likelihood tree of 33 SSU rDNA sequences and
1691 positions. Alexandrium tamarense was included as an outgroup. The best model, chosen by MrModel-Test2.3, was GTR+I+G. The support values shown were obtained by ML and BI. Only values larger than 50% (ML) and 0.50 (BI) are shown. A new sequence published in this study is displayed in bold (OP601437).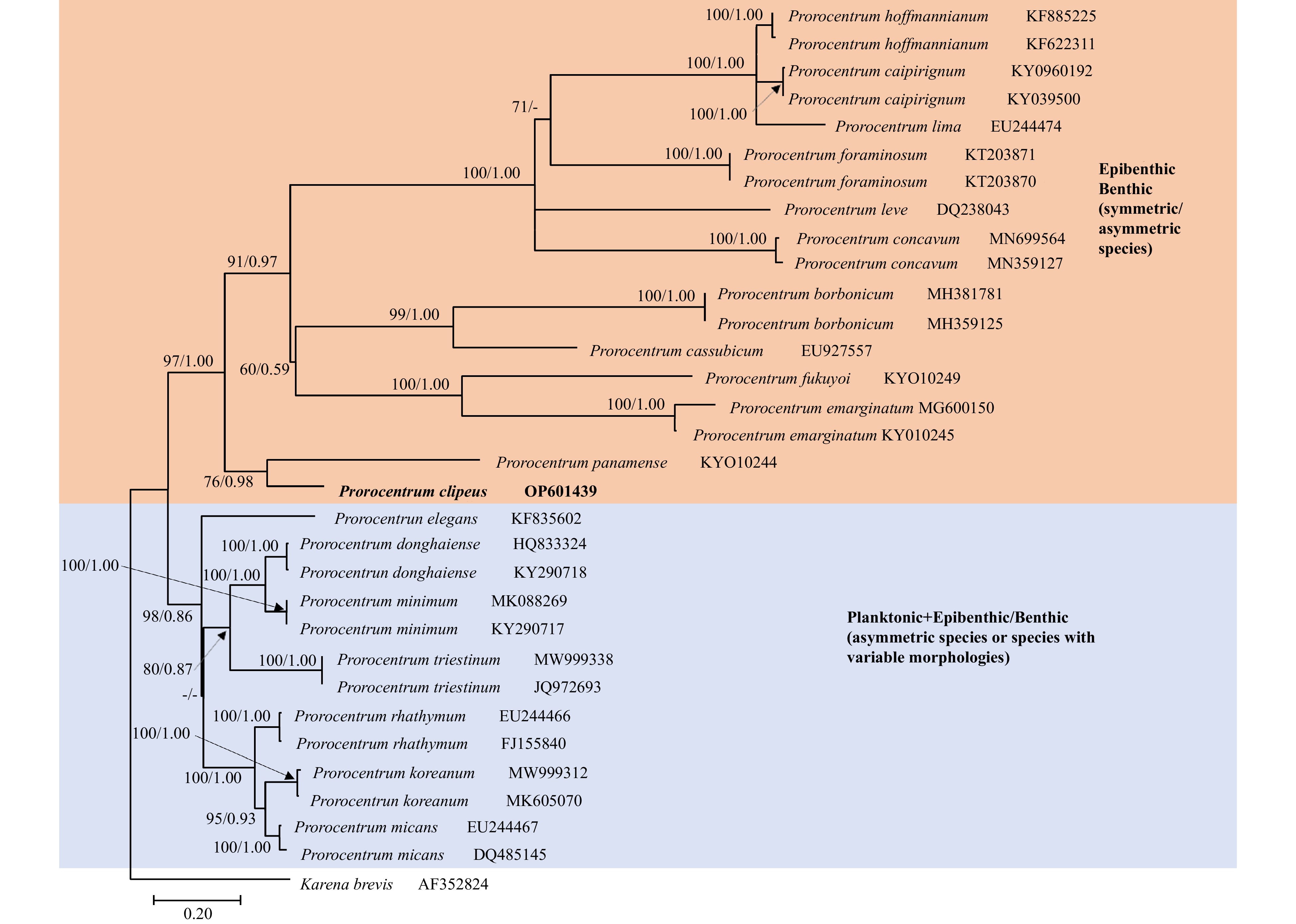
Figure 3. Maximum likelihood tree of 32 ITS1–5.8S–ITS2 sequences and 623 positions. Karenia brevis was included as an outgroup. The best model, chosen by MrModel-Test2.3, was GTR+I+G. The support values shown were obtained by ML and BI. Only values larger than 50% (ML) and 0.50 (BI) are shown. A new sequence published in this study is displayed in bold (OP601439).

Figure 4. Maximum likelihood tree of 37 LSU rDNA sequences and 697 positions. Alexandrium tamarense was included as an outgroup. The best model, chosen by MrModel-Test2.3, was GTR+I+G. The support values shown were obtained by ML and BI. Only values larger than 50% (ML) and 0.50 (BI) are shown. A new sequence published in this study is displayed in bold (OP601441).

Figure 5. Extracted ion chromatograms (EICs) of the procedure blank; P. clipeus DF128 intracellular and extracellular extracts; and OA, DTX1 and DTX2 standards (100 ng/mL) in negative ESI mode using a Sciex QTRAP 5500 system. cps: count per second.

Figure 6. The EICs of the procedural blank, P. clipeus DF128 intracellular and extracellular extracts, and standard PTX2 (100 ng/mL) in positive ESI mode were determined using a Sciex QTRAP 5500 system. cps: count per second.

Figure 7. Enhanced molecular networks obtained from the positive ion mode (a) and negative ion mode (b) mass spectra using MolNetEnhancer showing different molecular families/clusters of the pooled metabolites in the extracts of P. clipeus DF128. The node colors represent the classes of putatively annotated metabolites matched in the GNPS libraries. Single nodes indicate the absence of MS/MS fragments shared with any other compound.

Figure 8. Global occurrence of P. clipeus (data from this paper indicated by double circle marker; data from published literature and OBIS from 2000 to 2019 in black). The figure was created using Ocean Data View, version 5.7.1 (Schlitzer, 2023).
Table 1. Primer sequences used to amplify the SSU, ITS and LSU rDNA regions in Prorocentrum species
Name Target sequence Direction Sequence (5′ to 3′) Reference 18F23 SSU forward GGTTGATCCTGCCAGTAG Olmos-Soto et al. (2002) 18R1780 SSU reverse GTTCACCTACGGAAACCTTG Fu et al. (2008) SR4-F 548-566 SSU forward AGGGCAAGTCTGGTGCCAG Hong et al. (2008) SR5kawR 630-611 SSU reverse ACTACGAGCTTTTTAACCGC Hong et al. (2008) SR6-F 891-910 SSU forward GTCAGAGGTGAAATTCTTGG Hong et al. (2008) SR7-R 951-932 SSU reverse TCCTTGGCAAATGCTTTCGC Hong et al. (2008) SR9-R 1286 -1267 SSU reverse AACTAAGAACGGCCATGCAC Hong et al. (2008) ITS1-F ITS1-5.8S-ITS2 forward TCCGTAGGTGAACCTGCGG White et al. (1990) ITS4-R ITS1-5.8S-ITS2 reverse TCCTCCGCTTATTGATATGC White et al. (1990) DIR LSU forward ACCCGCTGAATTTAAGCATA Scholin et al. (1994) D2C LSU reverse CCTTGGTCCGTGTTTCAAGA Scholin et al. (1994)  下载: 导出CSV
下载: 导出CSV
Table 2. Mass spectrometric parameters and retention times of the OA, DTX1, DTX2 and PTX2 standards
Compound Molecular
formulaPrecursor
ion typePrecursor
ion (m/z)Fragment
ion (m/z)DP/V EP/V CE/eV CXP/V Retention
time/minOA C44H68O13 [M − H] − 803.5 255.2* −70 −10 −60 −12 7.45 113.1^ −70 −10 −75 −12 7.45 DTX1 C45H70O13 [M − H] − 817.5 255.2* −110 −10 −68 −12 8.57 113.1^ −110 −10 −94 −12 8.57 DTX2 C44H68O13 [M − H] − 803.5 255.2* −70 −10 −60 −12 7.72 113.1^ −70 −10 −75 −12 7.72 PTX2 C47H70O14 [M + NH4] + 876.4 823.5* 150 10 27 15 7.94 805.5^ 150 10 35 15 7.94 Note: *, quantificationion; ^, confirmationion.
下载: 导出CSV
Table 4. LSU rDNA sequence differences (above the diagonal line) and similarities (below the diagonal line) among P. clipeus, P. compressum and P. tsawwassenense based on a total of 697 positions
Species GenBank No./Strains Origin DF128 IFR459 IFR470 PCPA01 IFR456 P. clipeus OP601441/DF128 China − 39 39 95 140 P. clipeus JX912174/IFR459 France 94.4% − 0 104 121 P. clipeus JX912175/IFR470 France 94.4% 100% − 104 121 P. compressum AY259169/PCPA01 Australia 86.37% 85.08% 85.08% − 117 P. tsawwassenense JX912182/IFR456 France 79.91% 82.64% 82.64% 83.21% − Note: − represents no data.
下载: 导出CSV
Table 3. Comparison of the morphological features of Prorocentrum clipeus and similar benthic Prorocentrum species
Characteristic P. clipeus (this study) P. clipeus1 P. clipeus2 P. compressum5 P. tsawwassenense7 P. panamense8 Cell shape nearly round nearly round nearly circular ovate to rotundate oval heart-shaped Cell size Length/µm 37.8−41.3 54−55 37−55 (37−44)3; (30−35)4 30−50 (35−50)6 40−55 46−52 (52.3−55.6)9 Width/µm 35.7−39.7 50−52.5 32−36.54 C. 25(20−30)6 30−47.5 43−46 (48.3−50.7)9 L/W 1.00−1.09 1.05−1.08 ? ? ? 1.06−1.139 Periflagellar area Shape wide U-shaped wide arc-shaped wide U-shaped (arc) slight depression wide U-shaped linear Collar on left plate Yes Yes Yes No? Yes No Ridge on right plate Yes Yes Yes? No No No Wing-shaped spine No No No Yes “Yes” protrusions No Protrusions Only one Yes “Yes” apical spine Yes ? No More than one ? 5 (6)2 No Platelet list(s) No No No ? No No No. of platelets 9 10 9 ? 7-9 (8-10)2 ? 99 Flagellar pore Yes Yes Yes ? Yes Yes Accessory pore Yes Yes? Yes? ? Yes2 Yes Theca ornamentation smooth smooth smooth foveate6 smooth areolate (reticulate-foveate)2, 9 Pore pattern Yes, some cell visible pore pattern or radial rows No, scattered No visible pore pattern or radial rows of small pores (radial rows)3 No, scattered (rows of pores)6 Radial rows
Two apical rowsNo, scattered on valves, mostly around Platelet pores Yes Yes Yes? ? No No Marginal pores No No ? ? Yes ? No5 Plate center Devoid Devoid ? Yes? Devoid ? (Devoid in some cells)2 Large pores/µm No No ? ? 0.3–0.5 No Small pores/µm Approximately 0.15 µm in diameter Approximately 0.12 µm in diameter ? ? 0.09–0.17 0.15 Intercalary band Transverse striation Smooth ? (horiz. str.)3 ? Transverse and horizontal striation Transversally striated Pyrenoid Yes (LSCM) Probably yes (LM) ? ? Yes (TEM) Yes Nucleus (shape and position) Large kidney-shaped, Posterior Large kidney-shaped, Posterior Kidney-shaped, Posterior ? Round to oval, Posterior U-shaped, Posterior Note: In the above table, the list of morphological features was made based on Hoppenrath et al. (2013). It is listed here with some modifications, where notifications in the table indicate: ? = no data available; $\cdots $? = not mentioned in the text, inferred from images. Literature: 1Hoppenrath (2000); 2Hoppenrath et al. (2013); 3Murray (2003); 4Shah et al. (2013); 5Dodge (1975); 6Gul and Saifullah (2011); 7Hoppenrath and Leander (2008); 8Grzebyk et al. (1998); 9Luo et al. (2017).
下载: 导出CSV
-
Andersen R A. 2005. Algal Culturing Techniques. Oxford: Elsevier Academic Press, 578 Arteaga-Sogamoso E, Rodríguez F, Amato A, et al. 2023. Morphology and phylogeny of Prorocentrum porosum sp. nov. (Dinophyceae): a new benthic toxic dinoflagellate from the Atlantic and Pacific Oceans. Harmful Algae, 121: 102356, doi: 10.1016/j.hal.2022.102356 Blanco J, Moroño Á, Fernández M L. 2005. Toxic episodes in shellfish, produced by lipophilic phycotoxins: an overview. Galician: Revista Galega dos Recursos Mariños Galician Journal of Marine Resources, 1: 1–70 Boopathi T, Faria D G, Cheon J Y, et al. 2015. Implications of high molecular divergence of nuclear rRNA and phylogenetic structure for the dinoflagellate Prorocentrum (Dinophyceae, Prorocentrales). Journal of Eukaryotic Microbiology, 62(4): 519–531, doi: 10.1111/jeu.12206 Borsato G T, Salgueiro F, De’Carli G A L, et al. 2023. Taxonomy and abundance of epibenthic Prorocentrum (Dinophyceae) species from the tropical and subtropical Southwest Atlantic Ocean including a review of their global diversity and distribution. Harmful Algae, 127: 102470, doi: 10.1016/j.hal.2023.102470 Chomérat N, Bilien G, Zentz F. 2019. A taxonomical study of benthic Prorocentrum species (Prorocentrales, Dinophyceae) from Anse Dufour (Martinique Island, eastern Caribbean Sea). Marine Biodiversity, 49(3): 1299–1319, doi: 10.1007/s12526-018-0913-6 Chomérat N, Saburova M, Bilien G, et al. 2012. Prorocentrum bimaculatum sp. nov. (Dinophyceae, Prorocentrales), a new benthic dinoflagellate species from Kuwait (Arabian Gulf). Journal of Phycology, 48(1): 211–221, doi: 10.1111/j.1529-8817.2011.01102.x Chomérat N, Sellos D Y, Zentz F, et al. 2010. Morphology and molecular phylogeny of Prorocentrum consutum sp. nov. (Dinophyceae), a new benthic dinoflagellate from south brittany (northwestern France). Journal of Phycology, 46(1): 183–194, doi: 10.1111/j.1529-8817.2009.00774.x Chomérat N, Zentz F, Boulben S, et al. 2011. Prorocentrum glenanicum sp. nov. and Prorocentrum pseudopanamense sp. nov. (Prorocentrales, Dinophyceae), two new benthic dinoflagellate species from South Brittany (northwestern France). Phycologia, 50(2): 202–214, doi: 10.2216/10-12.1 Dodge J D. 1975. The Prorocentrales (Dinophyceae). II. Revision of the taxonomy within the genus Prorocentrum. Botanical Journal of the Linnean Society, 71(2): 103–125, doi: 10.1111/j.1095-8339.1975.tb02449.x Ehrenberg C. 1834. Dritter beitrag zur erkenntniss grosser organisation in der richtung des kleinsten raumes. Berlin: Abhandlungen der Königlichen Akademie der Wissenschaften zu, 1833: 145–336 Ernst M, Kang K B, Caraballo-Rodríguez A M, et al. 2019. MolNetEnhancer: enhanced molecular networks by integrating metabolome mining and annotation tools. Metabolites, 9(7): 144, doi: 10.3390/metabo9070144 European Food Safety Authority. 2008. Marine biotoxins in shellfish‐okadaic acid and analogues‐scientific opinion of the panel on contaminants in the food chain. European Food Safety Authority Journal, 6(1): 589, doi: 10.2903/j.efsa.2008.589 Faust M A, Vandersea M W, Kibler S R, et al. 2008. Prorocentrum levis, a new benthic species (Dinophyceae) from a mangrove island, Twin Cays, Belize. Journal of Phycology, 44(1): 232–240, doi: 10.1111/j.1529-8817.2007.00450.x Freudenthal A, Jijina J. 1988. Potential hazards of Dinophysis to consumers and shellfisheries. Journal of Shellfish Research, 7(1): 157–158 Fu X H, Meng F L, Hu Y, et al. 2008. Candida albicans, a distinctive fungal model for cellular aging study. Aging Cell, 7(5): 746–757, doi: 10.1111/j.1474-9726.2008.00424.x Glibert P M, Burkholder J M, Kana T M. 2012. Recent insights about relationships between nutrient availability, forms, and stoichiometry, and the distribution, ecophysiology, and food web effects of pelagic and benthic Prorocentrum species. Harmful Algae, 14: 231–259, doi: 10.1016/j.hal.2011.10.023 Gómez F, Gourvil P, Li T C, et al. 2023. Molecular phylogeny of the spiny‐surfaced species of the dinoflagellate Prorocentrum with the description of P. thermophilum sp. nov. and P. criophilum sp. nov. (Prorocentrales, Dinophyceae). Journal of Phycology, 59(1): 70–86, doi: 10.1111/jpy.13298 Grzebyk D, Sako Y, Berland B. 1998. Phylogenetic analysis of nine species of Prorocentrum (Dinophyceae) inferred from 18S ribosomal DNA sequences, morphological comparisons, and description of Prorocentrum panamensis, sp. nov. Journal of Phycology, 34(6): 1055–1068, doi: 10.1046/j.1529-8817.1998.341055.x Gul S, Saifullah S M. 2011. The dinoflagellate genus Prorocentrum (Prorocentrales, Prorocentraceae) from the north Arabian sea. Pakistan Journal of Botany, 43(6): 3061–3065 Gurevich A, Mikheenko A, Shlemov A, et al. 2018. Increased diversity of peptidic natural products revealed by modification-tolerant database search of mass spectra. Nature Microbiology, 3(3): 319–327, doi: 10.1038/s41564-017-0094-2 Hall T A. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41: 95–98 Han M S, Wang Pengbin, Kim J H, et al. 2016. Morphological and molecular phylogenetic position of Prorocentrum micans sensu stricto and description of Prorocentrum koreanum sp. nov. from southern coastal waters in Korea and Japan. Protist, 167(1): 32–50, doi: 10.1016/j.protis.2015.12.001 Henrichs D W, Scott P S, Steidinger K A, et al. 2013. Morphology and phylogeny of Prorocentrum texanum sp. nov. (Dinophyceae): a new toxic dinoflagellate from the Gulf of Mexico coastal waters exhibiting two distinct morphologies. Journal of Phycology, 49(1): 143–155, doi: 10.1111/jpy.12030 Holman J D, Tabb D L, Mallick P. 2014. Employing ProteoWizard to convert raw mass spectrometry data. Current Protocols in Bioinformatics, 46(1): 13.24. 1–13.24. 9, doi: 10.1002/0471250953.bi1324s46 Hong D D, Thu N T H, Nam H S, et al. 2008. The phylogenetic tree of Alexandrium, Prorocentrum and Pseudo-nitzschia of harmful and toxic algae in vietnam coastal waters based on sequences of 18s rDNA, ITS1-5.8S-ITS2 gene fragments and single cell-PCR method. Marine Research in Indonesia, 32(2): 203–218, doi: 10.14203/mri.v32i2.456 Hoppenrath M. 2000. A new marine sand-dwelling Prorocentrum species, P. clipeus sp. nov. (Dinophyceae, Prorocentrales) from Helgoland, German Bight, North Sea. European Journal of Protistology, 36(1): 29–33, doi: 10.1016/S0932-4739(00)80019-X Hoppenrath M, Chomérat N, Horiguchi T, et al. 2013. Taxonomy and phylogeny of the benthic Prorocentrum species (Dinophyceae)—a proposal and review. Harmful Algae, 27: 1–28, doi: 10.1016/j.hal.2013.03.006 Hoppenrath M, Leander B S. 2008. Morphology and molecular phylogeny of a new marine sand-dwelling Prorocentrum species, P. tsawwassenense (Dinophyceae, Prorocentrales), from British Columbia, Canada. Journal of Phycology, 44(2): 451–466, doi: 10.1111/j.1529-8817.2008.00483.x Hoppenrath M, Murray S A, Chomérat N, et al. 2014. Marine Benthic Dinoflagellates-Unveiling Their Worldwide Biodiversity. Stuttgart: Senckenberg, 137–138 Keller M D, Selvin R C, Claus W, et al. 1987. Media for the culture of oceanic ultraphytoplankton. Journal of Phycology, 23(4): 633–638, doi: 10.1111/j.1529-8817.1987.tb04217.x Jackson A E, Marr J C, Mclachlan J L. 1993. The production of diarrhetic shellfish toxins by an isolate of Prorocentrum lima from Nova Soctia, Canada. In: Smayda TJ, Shimizu Y, eds. Toxic Phytoplankton Blooms in the Sea. Newport: Elsevier, 513–518 Lim Z F, Luo Zhaohe, Lee L K, et al. 2019. Taxonomy and toxicity of Prorocentrum from Perhentian Islands (Malaysia), with a description of a non-toxigenic species Prorocentrum malayense sp. nov. (Dinophyceae). Harmful Algae, 83: 95–108, doi: 10.1016/j.hal.2019.01.007 Luo Zhaohe, Zhang Hua, Krock B, et al. 2017. Morphology, molecular phylogeny and okadaic acid production of epibenthic Prorocentrum (Dinophyceae) species from the northern South China Sea. Algal Research, 22: 14–30, doi: 10.1016/j.algal.2016.11.020 Mohimani H, Gurevich A, Shlemov A, et al. 2018. Dereplication of microbial metabolites through database search of mass spectra. Nature Communications, 9(1): 4035, doi: 10.1038/s41467-018-06082-8 Murray S. 2003. Diversity and phylogenetics of sand-dwelling dinoflagellates from southern Australia [dissertation]. Sydney: University of Sydney Murray S, Nagahama Y, Fukuyo Y. 2007. Phylogenetic study of benthic, spine-bearing prorocentroids, including Prorocentrum fukuyoi sp. nov. Phycological Research, 55(2): 91–102, doi: 10.1111/j.1440-1835.2007.00452.x Nascimento S M, Mendes M C Q, Menezes M, et al. 2017. Morphology and phylogeny of Prorocentrum caipirignum sp. nov. (Dinophyceae), a new tropical toxic benthic dinoflagellate. Harmful Algae, 70: 73–89, doi: 10.1016/j.hal.2017.11.001 Nothias L F, Petras D, Schmid R, et al. 2020. Feature-based molecular networking in the GNPS analysis environment. Nature Methods, 17(9): 905–908, doi: 10.1038/s41592-020-0933-6 Olmos-Soto J, Paniagua-Michel J, Contreras R, et al. 2002. Molecular identification of β-carotene hyper-producing strains of Dunaliella from saline environments using species-specific oligonucleotides. Biotechnology Letters, 24(5): 365–369, doi: 10.1023/A:1014516920887 Prabowo D A, Agusti S. 2019. Free-living dinoflagellates of the central Red Sea, Saudi Arabia: Variability, new records and potentially harmful species. Marine Pollution Bulletin, 141: 629–648, doi: 10.1016/j.marpolbul.2019.03.012 Rein K S, Snyder R V. 2006. The biosynthesis of polyketide metabolites by dinoflagellates. Advances in Applied Microbiology, 59: 93–125, doi: 10.1016/S0065-2164(06)59004-5 Ronquist F, Teslenko M, Van Der Mark P, et al. 2012. MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology, 61(3): 539–542, doi: 10.1093/sysbio/sys029 Schlitzer R. 2023. Ocean data view. http://odv.awi.de Schmid R, Heuckeroth S, Korf A, et al. 2023. Integrative analysis of multimodal mass spectrometry data in MZmine 3. Nature Biotechnology, 41(4): 447–449, doi: 10.1038/s41587-023-01690-2 Scholin C A, Herzog M, Sogin M, et al. 1994. Identification of group‐and strain‐specific genetic markers for globally distributed Alexandrium (Dinophyceae). II. Sequence analysis of a fragment of the LSU rRNA gene. Journal of Phycology, 30(6): 999–1011, doi: 10.1111/j.0022-3646.1994.00999.x Shah M M R, An S J, Lee J B. 2013. Presence of benthic dinoflagellates around coastal waters of Jeju Island including newly recorded species. Journal of Ecology and Environment, 36(4): 347–370, doi: 10.5141/ecoenv.2013.347 Shannon P, Markiel A, Ozier O, et al. 2003. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Research, 13(11): 2498–2504, doi: 10.1101/gr.1239303 Swofford D L, 2002. PAUP: phylogenetic analysis using parsimony, version 4.0 b10. Sinauer Associates, Sunderland, MA Ten-Hage L, Turquet J, Quod J P, et al. 2000. Prorocentrum borbonicum sp. nov. (Dinophyceae), a new toxic benthic dinoflagellate from the southwestern Indian Ocean. Phycologia, 39(4): 296–301, doi: 10.2216/i0031-8884-39-4-296.1 Thompson J D, Higgins D G, Gibson T J. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic acids research, 22(22): 4673–4680, doi: 10.1093/nar/22.22.4673 Tillmann U, Gottschling M, Wietkamp S, et al. 2023a. Morphological and phylogenetic characterisation of Prorocentrum spinulentum, sp. nov. (Prorocentrales, Dinophyceae), a small spiny species from the North Atlantic. Microorganisms, 11(2): 271, doi: 10.3390/microorganisms11020271 Tillmann U, Hoppenrath M, Gottschling M. 2019. Reliable determination of Prorocentrum micans Ehrenb. (Prorocentrales, Dinophyceae) based on newly collected material from the type locality. European Journal of Phycology, 54(3): 417–431, doi: 10.1080/09670262.2019.1579925 Tillmann U, Wietkamp S, Gottschling M, et al. 2023b. Prorocentrum pervagatum sp. nov. (Prorocentrales, Dinophyceae): a new, small, planktonic species with a global distribution. Phycological Research, 71(1): 56–71, doi: 10.1111/pre.12502 Verma A, Kazandjian A, Sarowar C, et al. 2019. Morphology and phylogenetics of benthic Prorocentrum Species (Dinophyceae) from tropical northwestern Australia. Toxins, 11(10): 571, doi: 10.3390/toxins11100571 Wan Xiukun, Yao Ge, Liu Yanli, et al. 2019. Research progress in the biosynthetic mechanisms of marine polyether toxins. Marine Drugs, 17(10): 594, doi: 10.3390/md17100594 Wu Yixuan, Huang Shuning, Krock B, et al. 2022. Cryptic speciation of benthic Prorocentrum (Dinophyceae) species and their potential as ecological indicators. Journal of Sea Research, 190: 102304, doi: 10.1016/j.seares.2022.102304 White T J, Bruns T, Lee S, et al. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis M A, Gelfand D H, Sninsky J J, et al, eds. PCR Protocols: A Guide to Methods and Applications. New York: Academic Press, 315–322, doi: 10.1016/B978-0-12-372180-8.50042-1 Xie Hang, Zou Jian, Zheng Chengzhi, et al. 2022. Biodiversity and distribution of benthic dinoflagellates in tropical Zhongsha Islands, South China Sea. Journal of Oceanology and Limnology, 40(6): 2120–2145, doi: 10.1007/s00343-022-1322-z Yasumoto T, Murata M, Oshima Y, et al. 1985. Diarrhetic shellfish toxins. Tetrahedron, 41(6): 1019–1025, doi: 10.1016/S0040-4020(01)96469-5 Zhang Hua, Li Yang, Cen Jingyi, et al. 2015. Morphotypes of Prorocentrum lima (Dinophyceae) from Hainan island, South China Sea: morphological and molecular characterization. Phycologia, 54(5): 503–516, doi: 10.2216/15-8.1 Zou Jian, Li Qun, Liu Hui, et al. 2022. Taxonomy and toxin profile of harmful benthic Prorocentrum (Dinophyceae) species from the Xisha Islands, South China Sea. Journal of Oceanology and Limnology, 40(3): 1171–1190, doi: 10.1007/s00343-021-1045-6 Zou Jian, Li Qun, Lu Songhui, et al. 2020. The first benthic harmful dinoflagellate bloom in China: morphology and toxicology of Prorocentrum concavum. Marine Pollution Bulletin, 158: 111313, doi: 10.1016/j.marpolbul.2020.111313 -

 DownLoad:
DownLoad:


 点击查看大图
点击查看大图
计量
- 文章访问数: 326
- HTML全文浏览量: 140
- PDF下载量: 70
- 被引次数: 0

