Ze Meng, Lei Zhou, Baosheng Li, Jianhuang Qin, Juncheng Xie. Erratum to: Acta Oceanologica Sinica (2022) 41(10): 119–130DOI: 10.1007/s13131-022-2023-3The atmospheric hinder for intraseasonal sea-air interaction over the Bay of Bengal during Indian summer monsoon in CMIP6[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-022-2131-0
Citation:
Vicent Michael Shija, Kwaku Amoah, Yi Li, Yong Zhong, Xiaopiao Zhong, Jia Cai. In vitro selection, assessment, and identification of potential probiotic features of the Bacillus strains isolated from the intestine of Nile tilapia (Oreochromis niloticus)[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-024-2411-y
Ze Meng, Lei Zhou, Baosheng Li, Jianhuang Qin, Juncheng Xie. Erratum to: Acta Oceanologica Sinica (2022) 41(10): 119–130DOI: 10.1007/s13131-022-2023-3The atmospheric hinder for intraseasonal sea-air interaction over the Bay of Bengal during Indian summer monsoon in CMIP6[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-022-2131-0
Citation:
Vicent Michael Shija, Kwaku Amoah, Yi Li, Yong Zhong, Xiaopiao Zhong, Jia Cai. In vitro selection, assessment, and identification of potential probiotic features of the Bacillus strains isolated from the intestine of Nile tilapia (Oreochromis niloticus)[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-024-2411-y
In vitro selection, assessment, and identification of potential probiotic features of the Bacillus strains isolated from the intestine of Nile tilapia (Oreochromis niloticus)
College of Fishery, Guangdong Ocean University, Zhanjiang 524088, China
2.
Guangdong Provincial Key Laboratory of Aquatic Animal Disease Control and Healthy Culture, Zhanjiang 524088, China
3.
Guangdong Key Laboratory of Control for Diseases of Aquatic Economic Animals, Zhanjiang 524088, China
4.
Southern Marine Science and Engineering Guangdong Laboratory, Zhanjiang 524002, China
5.
Guangxi Key Laboratory for Marine Natural Products and Combinational Biosynthesis Chemistry, Nanning 530200, China
6.
Guangxi Beibu Gulf Marine Research Centre, Guangxi Academy of Sciences, Nanning 530007, China
Funds:
The National Natural Science Foundation of China under contracts Nos U20A2065, 32073006, and 32002426; the Natural Science Foundation of Guangxi under contract No. 2020GXNSFAA297243; the Natural Science Foundation of Guangdong Province under contract No. 2021B0202040002; the Science and Technology Plan of Guangdong Province under contract No. 2023B0202010016; the Youth Science and Technology Innovation Talent of Guangdong TeZhi Plan Talent under contract No. 2023TQ07A888; the Science and Technology Planning Project of Zhanjiang under contract Nos 2022A01012 and 2022A01006.
In this study, we aimed to isolate potential Bacillus strains from the digestive tract of Nile tilapia (Oreochromis niloticus). Through morphological, biochemical, 16S rDNA sequencing, and evolutionary relationship assessments, we identified the isolated species as Bacillus amyloliquefaciens AV5 (OR647358), B. subtilis AV7 (LC781790), and B. velezensis AV50 (OR647359), designated as AV5, AV7, and AV50 strains, respectively. We evaluated their probiotic potential, including tolerance to high bile salt concentrations, low pH levels, and high temperatures, as well as their adhesion abilities (auto-aggregation and cell-surface hydrophobicity), antimicrobial activity, biosafety, compatibility, hemolytic activity, and antibiotic susceptibility. AV5, AV7, and AV50 strains exhibited γ-hemolytic activity and resistance to low pH (1) and high bile salt concentrations (0.5%). They demonstrated higher viability after exposure to elevated temperatures (80℃, 90℃, and 100℃) and increased cell surface hydrophobicity and auto-aggregation. These strains showed positive compatibility, indicating their potential for multispecies use. Strains were susceptible to various antibiotics, showing susceptibility to 17 out of 20 tested antibiotics, and displayed significant antimicrobial activity against Vibrio alginolyticus, V. harveyi, Streptococcus iniae, and S. agalactiae. Therefore, AV5, AV7, and AV50 are promising probiotic candidates for enhancing the growth and health of aquatic animals, particularly the Nile tilapia.
The use of antibiotics in aquaculture can lead to changes in the gut flora and bacterial resistance, posing a threat to aquatic health (Dawood et al., 2018; Hoseinifar et al., 2020). Alternative approaches, such as probiotics, medicinal plants, vaccines, para-probiotics, and prebiotics have been explored (Choudhury and Kamilya, 2019; Widanarni et al., 2022). Probiotics possess unique traits that enable them to thrive in the gastrointestinal tract, including resistance to gastric acidity and bile salts, tolerance to temperature fluctuations, production of antimicrobial compounds, and the ability to adhere to intestinal surfaces (Ringø et al., 2020).
Moreover, probiotic strains must adhere to biosafety standards, including non-hemolytic activity and antibiotic susceptibility (Ringø et al., 2020). Probiotics show great potential for aquaculture by offering a range of benefits compared to other preventive methods (Hlordzi et al., 2020). Bacillus sp., particularly probiotics Bacillus (e.g., B. amyloliquefaciens, B. velezensis, B. subtilis), exhibit distinct advantages such as the production of non-pathogenic substances, improvement of water quality, and resilience under harsh conditions due to spore formation (Kavitha et al., 2018).
Several Bacillus species such as B. velezensis, B. amyloliquefaciens, and B. subtilis are significant due to their capacity to synthesize metabolites or antibiotics that have antagonistic impacts against pathogenic microbes (Kuebutornye et al., 2020). They have been utilized in the medical and pharmaceutical sectors to manage various diseases in animals, humans, and plants as biological control agents because of their capacity to produce a diverse array of metabolites with antibacterial properties (Amenyogbe et al., 2024). Bacillus species are known to boost fish feed consumption, digestive enzyme activity, antioxidant enzyme activity, and the expression of immune-related and stress-related genes to enhance the capacity of fish to resist pathogenic microorganisms (Amenyogbe et al., 2024; Amenyogbe, 2023).
Numerous studies have highlighted the beneficial effects of Bacillus probiotics in Nile tilapia farming (Abarike et al., 2018; El-Dakar et al., 2023; Vijayaram et al., 2024; Abarike et al., 2018). Aly et al. (2008) found that B. licheniformis Dahb1 and B. pumilus substantially impacted Nile tilapia's growth. The growth performance and intestinal morphology of Nile tilapia, O. niloticus, were enhanced by dietary supplementation with B. subtilis, B. licheniformis, and Bacillus pumilus, either in combination or in a dose-dependent manner (Elsabagh et al., 2018). Administration of B. subtilis to tilapia diets enhanced immune responses and particle engulfment (Shija et al., 2023).
The probiotic Bacillus species used in aquaculture are sourced from fish intestines, soil, water, organic matter, vertebrates, and commercial substrates. However, commercial probiotics often lack efficacy in aquaculture settings because of their non-fish origin, which hinders effective gut colonization (Vijayaram et al., 2024). Some studies have suggested that probiotics derived from fish intestines exhibit superior efficacy compared with those derived from other sources (Van Doan et al., 2018).
However, studies on the specific effects of individual or combined Bacillus strains on tilapia farming under varying environmental conditions remain limited. Therefore, this study aimed to isolate and evaluate novel Bacillus species from the intestine of Nile tilapia (O. niloticus) as potential probiotics.
2.
Materials and methods
2.1
Animal sample collection
A total of 50 wild healthy Nile tilapia (O. niloticus) of average weight 33.24±0.28 g showing no signs of infection (Abarike et al., 2018), were collected from a local fish lake at West Lake, Guangdong Ocean University, Zhanjiang City, Guangdong Province, China (21.2682°N and 110.3589°E) using a dip net. Live fish were transported to the laboratory in aerated bags for the immediate commencement of the experiment (Abarike et al., 2018).
2.2
Strain isolation and culture conditions
Healthy fish with a body weight of approximately 1600 g were anesthetized using high doses of ethyl 3-ammonium benzoate and tricaine sulfonate (Sigma-Aldrich, 150 mg/L MS-222), and then killed by being struck on the head. The cotton was soaked in 75% ethanol and then cleaned externally to remove any external bacteria from the fish bodies. Sterile pairs of scissors were used to dissect the fish gut. The intestines were removed and carefully stripped to remove digesta content before being washed three times with a physiological saline solution (PBS). An equal proportion of PBS was added to the weight of the intestine by volume. The contents were then homogenized using a 15 mL borosilicate glass tissue homogenizer under sterile conditions in ice to maintain their temperature. Gut homogenate (0.5 mL of gut homogenate was mixed with 5 mL PBS, serially diluted, and then 0.1 mL of the aliquot was spread on Luria-Bertani (LB) agar plates and incubated at 37℃ for 24 h. To obtain very pure colonies, single colonies were randomly selected and inoculated into the LB medium under the same culture conditions.
2.3
PCR, and strain identification
2.3.1
Amplification
Potential probiotic strains were characterized based on the morphological, and biochemical tests (compatibility test, pH determination, bile salt tolerance, antimicrobial tests resistance in high-temperature, biosafety test, antibiotic sensitivity test, auto-aggregation, hemolytic test hydrophobicity, and biofilm formation), and identified by molecular 16S rDNA from a bacterial suspension using universal bacterial primers 27F (AGAGTTTGATCCTGGCTCAG) and 1492R (GGTTACCTTGTTACGACTT) in a polymerase chain reaction (PCR). A total of 2 μL of each primer, 2 μL template of each isolate, 5 μL of 10× Extaq buffer, and 39 μL of double distilled water were used in the PCR reaction system (Kuebutornye et al., 2018). Distilled water was used as a negative control template, and Bacillus subtilis (MW548634) (previously isolated in our laboratory) was used as a positive control template (Amoah et al., 2021). PCR amplification was initiated by denaturation at 96℃ for 5 min, followed by 33 cycles of denaturation at 96℃ for 30 s, annealing at 55℃ for 45 s, and extension at 72℃ for 1 min 30 s. Amplification was performed by incubating the reaction mixture at 72℃ for 10 min. The ethidium bromide PCR products were analyzed by agarose gel (1%, w/v) electrophoresis under the following conditions: 80-150 V, 210 mA, and 20 min. The gel images were detected using a gel imager.
2.3.2
Cloning
The cloning vector, pMD19-T (Amp+), was purchased from TaKaRa Biotechnology (Dalian, China). Escherichia coli DH5α cells were maintained in our laboratory. Ampicillin stock solution (100 mg/mL) was prepared with sterile water and stored at -20℃. The working concentration was 100 µL/100 mL in the medium. The chloramphenicol stock solution was prepared using anhydrous ethanol at a working concentration of 25 µg/mL. A calcium chloride solution (0.1 mol/g) was prepared to prepare competent cells by dissolving 1.7 g of CaCl2·2H2O in 100 mL of sterile water, filtering through a 0.22 µm filter, and storing in a refrigerator at 4℃. Strains were identified according to modified methods described by Kuebutornye et al. (2018). Briefly, the purified PCR products (16S rDNA (4.5 µL) were ligated with pMD19-T (0.5 µL) vector at 16℃ for 2 h in a water bath machine and transformed into E. coli DH5α (33 µL) competent cells, and the transformants were selected using white screening. Plasmids containing the 16S rDNA were extracted and purified (Kuebutornye et al., 2018). Positive colonies were identified and confirmed by colony PCR.
2.3.3
Genotypic identification of the three isolates
The genotypic identification of the three isolates was performed by Sangon Biotech Co., Ltd. (Guangzhou, China) based on16S rDNA gene sequencing, which was subsequently compared with available sequences in the National Center for Biotechnology Information (NCBI) using the Basic Local Alignment Search Tool (BLASTn) program to confirm the bacterial species. A phylogenetic tree was generated by the neighbor-joining (NJ) method using the NCBI Distance Tree online tool. All the selected strains were stored in an equivalent mixed solution of 40% (v/v) glycerol and LB for future use.
2.4
Biochemical tests of isolated strains
As demonstrated in Tables 1 and 2, phylogenetic and genetic analyses of isolates are compared through morphological biochemical characterization tests. Commercial kits obtained from Guangdong Huankai Microbial Science and Technology Co. Ltd. (Guangdong, China) were used to analyze the biochemical properties of the selected probiotic strains. We confirmed the biochemical results based on the Bacillus cereus identification bar (HBIG07-1) acquired from Qingdao Hope Bio-Tech Co., Ltd. (Qingdao, China).
Table
1.
Morphological features of the isolates from the intestine of the Nile tilapia (Oreochromis niloticus)
Isolate
Color
Form
Surface
Size
Margin
Elevation
Texture
AV7
white
circular
dry
medium
entire
raised
rough
AV5
creamy white
irregular
moist
medium
undulate
umbonate
rough
AV50
white
irregular
dry
medium
undulate
umbonate
rough
Note: The three isolated strains, AV5, AV7, and AV50, exhibited identity sequence homology (99%) to B. amyloliquefaciens, B. subtilis, and B. velezensis respectively.
Table
2.
The biochemical features of the selected strains isolated from the intestine of the Nile tilapia (Oreochromis niloticus)
Tests performed
Biochemical feature
Tests performed
Biochemical feature
AV50
AV7
AV5
AV50
AV7
AV5
Mannitol
+
−
−
Simon’s citrate
+
−
+
Lactose fermentation
–
–
–
Starch hydrolysis
+
+
+
Gram staining
+
+
+
Biofilm production
–
–
–
Glucose
+
+
+
Spore formation
+
+
+
Rhamnose
+
+
+
Arginine dihydrolase
+
+
+
Nitrate reduction
+
+
+
Sorbitol
+
+
+
Lysozyme broth
–
+
–
Inositol
+
+
+
Methyl red
–
–
–
Adonitol
+
+
+
Urease
+
+
+
Haemolysis
$\gamma $
$\gamma $
$\gamma $
Gelatin liquefaction
–
–
–
Catalase
+
+
+
Hippuric acid
–
–
–
Vorges proskauer
–
–
–
Note: The three isolated strains, AV5, AV7, and AV50, exhibited identity sequence homology (99%) to B. amyloliquefaciens, B. subtilis, and B. velezensis respectively. Additionally, + donated a positive response, – a negative response, and γ no response.
2.5
Growth patterns of the selected isolates in Luria–Bertani broth
A single colony of a probiotic bacterial strain was selected from LB agar (pH 7.3) and cultured for 12 h in 5 mL of LB broth at 37℃. A total of 1 mL of the overnight culture was added to 100 mL of LB broth (pH 7.2) in 500-mL Erlenmeyer flasks. The flasks were then placed in a shaking incubator at 150 r/min and 37℃ for 24 h. Throughout the culture phase, bacterial growth was monitored every 2 h until it reached the specified 24-h time point. This was done using a spectrophotometer (EvolutionTM 220 UV-Visible Spectrophotometer, Thermo ScientificTM, USA) to measure the optical density (OD) at 600 nm (Xie et al., 2014). Finally, the growth curves of various strains were plotted.
2.6
Biosafety test
We evaluated the potential adverse effects associated with probiotic strains in healthy omnivorous Nile tilapia (O. niloticus). Juvenile Nile tilapia with an average weight of 19.5±2.5 g was obtained from the local fish farm in Zhanjiang, China. The fish were acclimatized for 14 d in aerated tanks under controlled conditions. The temperature of the water (28±0.5℃) was assessed throughout the experiment. During this period, the fish were fed a commercial diet (crude protein ≥ 34.0% crude fat ≥ 2.50%, crude fibre ≤ 8.00%, crude ash ≤ 15.0%, moisture content ≤ 11.0%, lysine ≥ 1.80%, calcium 0.5%-2.50%, total phosphorus 0.6%-2.0% and NaCl ≤ 2.00%) (Zhanjiang Aohua Feed Co. Ltd., Guangdong, China, Q/ZJHR 02-2022) twice daily (9:00-9:30 am and 5:30-6:00 pm) at a rate of 3% of average body weight. After acclimatization, 180 fish (28.7±1.2 g) were randomly assigned to nine aerated tanks, each containing 20 fish for 30 d. The fish were divided into three experimental groups (G1, G2, and G3), each with three replicates. Before the experimental treatments, the fish were starved for 24 h. To evaluate the harmful effects of probiotic bacteria isolates, three groups of ten fish, each weighing 48.7 ± 1.25 g, were administered intraperitoneal injections of 0.1 mL of the respective probiotic strains (AV5, AV7, and AV50). Control fish received an equivalent volume of sterile phosphate-buffered saline (PBS) at pH 7.0. Fish were maintained under previously described culture conditions (Geng et al., 2012). Clinical signs of the disease and mortality rates were monitored and recorded daily for 14 d.
2.7
Antibiotic sensitivity test
The disc diffusion method was employed to assess the susceptibility of isolates (AV50, AV7, and AV5) to a panel of 20 antibiotics, as presented in Table 3. The antibiotics tested were kanamycin, norfloxacin, doxycycline, ceftriaxone, chloramphenicol, erythromycin, clindamycin, furazolidone, ampicillin, penicillin, polymyxin B, vancomycin, sulfamethoxazole, ofloxacin, gentamicin, tetracycline, midecamycin, ciprofloxacin, piperacillin, and cefoperazone (Hangzhou Microbial Reagent Co. Ltd., Hangzhou, China). Briefly, 0.1 mL of cultured isolated probiotics, adjusted to the optical density (OD) corresponding to No. 0.5 on the McFarland standard (equivalent to 1.5×108 colony-forming units per milliliter), was spread on Mueller–Hinton agar plates (Beijing Land Bridge Tech. Co., Ltd.). Subsequently, antibiotic discs were carefully positioned on the agar surface and incubated at 37℃ for 24 h. Following incubation, the susceptibility of probiotics was determined by measuring the diameter of the inhibition zone (in mm), as previously described (Amoah et al., 2021).
Table
3.
Sensitivity of isolates (mm) to antibiotics
Antibiotic
Antibiotic content/ µg·disc-1
Isolate
AV5
AV7
AV50
Tetracycline
30
S
S
S
Erythromycin
15
S
S
S
Chloramphenicol
30
S
S
S
Polymyxin B
300
S
S
MS
Penicillin
10
S
S
S
Ciprofloxacin
5
S
S
S
Kanamycin
30
S
S
S
Ampicillin
10
S
S
S
Gentamicin
10
S
S
S
Furazolidone
300
S
R
S
Clindamycin
2
S
S
S
Ceftriaxone
30
S
S
S
Cefoperazone
75
S
S
S
Doxycycline
30
S
S
S
Vancomycin
30
S
S
S
Midecamycin
30
S
S
S
Ofloxacin
5
S
S
S
Sulfamethoxazole
27
MS
S
S
Piperacillin
100
S
S
S
Norfloxacin
10
S
S
S
Note: Isolate antibiotics sensitivity was evaluated according to the McFarland standard, incubated at 37℃ for 24 h, and the susceptibility of the isolates was determined by measuring the diameter of the inhibition zone (in millimeters). S, R, and MS donate susceptible, resistant, and moderately susceptible responses respectively. B. amyloliquefaciens (AV5), B. velezensis (AV7), and B. velezensis (AV50).
Isolate resistance to bile salt (0.3%–0.5%) was determined following the methodology performed by Argyri et al. (2013) with slight modifications. Cultured bacterial strains (AV50, AV7, and AV5) were harvested, washed twice with phosphate-buffered saline (PBS) at pH 7.0, and subsequently suspended in PBS solution (pH 7.3 containing bile salts at a concentration of 0.5% (BBI Life Sciences, Shanghai, China) after a 12-h incubation period. Viable colonies of surviving isolates were counted after incubation at 37℃ for 1–3 h. The percent survival of the Bacillus strains was calculated using the following formula: survival (%) = (Nt/N0) × 100, where Nt represents the viable cells surviving after incubation when the bacterial isolates (AV5, AV7, and AV50) were exposed to PBS with 0.3%-0.5% bile salts for 1 h to 3 h, and N0 denotes the surviving cells after incubation of selected strains in PBS without bile salts. Tolerance of the isolated strains to bile salt was assessed in triplicate.
2.9
Isolates’ resistance in high-temperature
Following the methodology performed by Guo et al. (2016), with minor adjustments, the resistive capacity of the isolated bacterial strains at various temperatures was assessed. Given the necessity of high-temperature processing in fish feed and other animal feed production, it is imperative to understand the survival of isolated bacteria under such harsh conditions. In this regard, overnight cultures of the isolates were washed twice with 40 mL of phosphate-buffered saline (PBS) at pH 7.4 and subsequently subjected to temperatures of 80℃, 90℃, and 100℃ using a Med-L-Hh 6 Electrothermal Thermostatic water-bath heater (Guangzhou Med Equipment Co. Ltd.) for 2 min, 5 min, and 10 min, respectively. Following heat treatment, an equal volume of Luria-Bertani (LB) broth was added to the isolates to assess their growth ability after treatment. Growth was monitored by measuring colony-forming units (CFU) after 12 h of incubation at 37℃. The high-temperature resistance tests were conducted in triplicate.
2.10
Isolates compatibility test
A compatibility test was carried out following the method outlined by Rajyalakshmi et al. (2016). Selected probiotic isolates were streaked vertically at 5 mm on Luria-Bertani (LB) agar, followed by perpendicular streaks of 10 mm. Compatibility was assessed based on the presence or absence of a zone of inhibition between the isolates after 24 h of incubation at 37℃.
2.11
Antimicrobial features
Vibrio harveyi, V. alginolyticus, S. agalactiae, and S. iniae were provided by the Guangdong Key Laboratory of Control for Diseases of Aquatic Economic Animals, Zhanjiang, China, for use in our laboratory (Amoah et al., 2021). The cross-streak method and agar well diffusion methods were employed to assess pathogenic bacteria against Bacillus strains isolated from Nile tilapia, following the protocol outlined by Lee et al. (2017). Briefly, 0.1 mL of each pathogenic bacteria was inoculated into 100 mL of Luria-Bertani (LB) broth in a 250-mL flask and cultured at 160×g for 16 h at 37℃. Subsequently, the bacterial cells were pelleted by centrifugation at 4 930×g for 10 min at 4℃, purified twice with phosphate-buffered saline (PBS), and their concentration was adjusted based on the absorbance at 600 nm. Using serial dilution and the spread plate method, actual doses ranging from 106 to 109 colony-forming units per milliliter (CFU/mL) were obtained from the supernatants. To determine the most suitable concentration for the experiment, calibrated doses with isolates were tested, and a concentration of 1×108 CFU/mL was selected. The pathogens were then evaluated against selected Bacillus strains using cross-streak and agar well diffusion methods in triplicate.
2.12
Hydrophobicity of isolated strains
The hydrophobicity of the selected strains was analyzed using the method described by Lee et al. (2017), with slight modifications. The strains were assessed based on their ability to adhere to organic solvents (hydrophobicity). An overnight bacterial culture of Bacillus strains was centrifuged at 9 500×g for 5 min, washed twice, and then 2 mL of PBS (pH 7.2) was added. The absorbance of the cell suspension at 600 nm was measured to determine hydrophobicity (%) and recorded as H0. Equal amounts of solvent (ethyl acetate, xylene, and chloroform) were mixed with the cell suspension for 4 min. The suspension was incubated at 27℃ for 40 min to separate the two phases. The aqueous phase was removed, measured at 600 nm, and recorded as Ht. The hydrophobicity (%) was determined as follows: hydrophobicity (%) = 1– (Ht/H0) ×100, where Ht is the absorbance at 600 nm and H0 is the absorbance before.
2.13
Auto-aggregation of isolated strains
Auto-aggregation assays were performed using a modified method of Song et al. (2023). In LB broth, selected isolates were cultured overnight at 37℃. Selected isolates were harvested at 9 000×g for 4 min, purified 2 times with PBS (pH 7.0), resuspended in supernatant, and mixed for 40 s. The absorbance was determined at 600 nm using a UV/visible spectrophotometer at 0 h, 1 h, 2 h, 3 h, 4 h, and 24 h. Auto-aggregation (%) was determined as follows: auto-aggregation (%) = (1–[Bt/B0]) × 100, where Bt is the absorbance at time t (h) being 1, 2, 3, 4 or 24, and B0 is the absorbance at t = 0.
2.14
Hemolytic test
Strains of Bacillus species were cultured on agar plates containing 7% sheep blood, and hemolytic activity tests were conducted on each of the three isolated strains. After incubation for 48 h at 37℃, the hemolytic zones were examined on the plates. The isolates were categorized according to their hemolysis patterns as α-, β-, and γ-hemolysis. The isolates that exhibited a green zone around the colony were noted as α-hemolysis, a clear zone was recorded as β-hemolysis and no changes on the agar for γ-hemolysis (Lee et al., 2017; Amenyogbe et al., 2021).
2.15
Optimal growth and pH of the isolated strains
The growth rate and pH were assessed according to the methodology described by Kavitha et al. (2018). In summary, fresh overnight cultures of the three bacterial isolates were introduced into LB broth at concentrations ranging from 1 to 10. The pH of the broth was adjusted using acetic acid (99%) and 5 mol/L NaOH. Infected broths were then placed in an incubator at 37℃ for 24 h. Subsequently, a spectrophotometer (EvolutionTM 220 UV-Visible Spectrophotometer, Thermo Scientific TM, USA) was used to monitor the growth of the inoculated broths relative to the uninoculated broth at a wavelength of 600 nm.
2.16
Biofilm formation test
Biofilm production analysis was conducted as described by Kavitha et al. (2018). Selected isolates (AV5, AV7, and AV50) were streaked on Mueller-Hinton agar supplemented with 0.8 g/L Congo red dye (Shanghai Macklin Biochemical Co., Ltd.) and incubated for 48 h at 37℃. The appearance of black colonies with dry crystals indicated biofilm production, whereas the presence of red colonies indicated that the strains were incapable of forming biofilms.
2.17
Statistical analysis
For all experiments, we performed a one-way analysis of variance (ANOVA) using the Statistical Package for Social Sciences (SPSS) software (IBM SPSS v20.0, Inc., 2010, Chicago, USA). The difference between means was determined using Tukey's honestly significant difference (HSD) test. A significant difference (p<0.05) was considered statistically significant, and the results were expressed as the mean ± standard deviation (SD).
3.
Results
3.1
Identification of isolated probiotic Bacillus strains
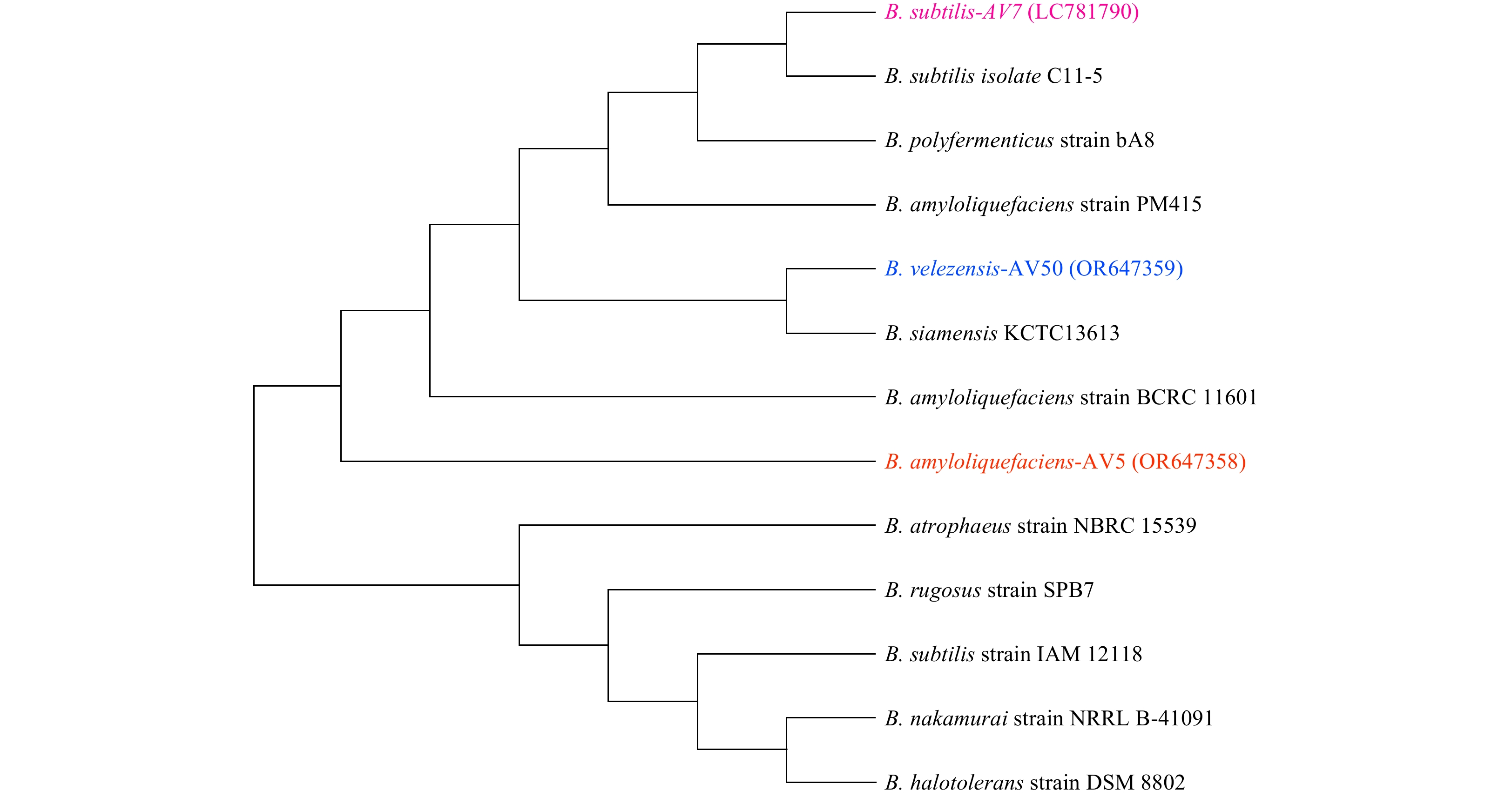
Tables 1 and 2 present the morphological and biochemical characteristics of the isolates, respectively. AV5, AV7, and AV50 showed 99% sequence homology with those of B. amyloliquefaciens, B. subtilis, and B. velezensis, respectively. The isolated strains retained the crystal violet stain, appearing as purple rods, confirming their gram-positive nature. Phylogenetic analysis (Fig. 1) indicated that isolates AV5, AV7, and AV50 exhibited the greatest genetic similarity to B. amyloliquefaciens, B. subtilis, and B. velezensis, respectively. The nucleotide sequences of the three strains have been deposited in NCBI GenBank with accession numbers: (OR647358) B. amyloliquefaciens AV5, (LC781790) B. subtilis AV7, and (OR647359) B. velezensis AV50.
Figure
1.
A phylogenetic tree based on 16SrDNA sequences generated by the neighbor-joining method (1 000 bootstraps) using the NCBI distance tree. This figure illustrates the relationship between closely related Bacillus species and selected Bacillus strains (AV5, AV7, and AV50).
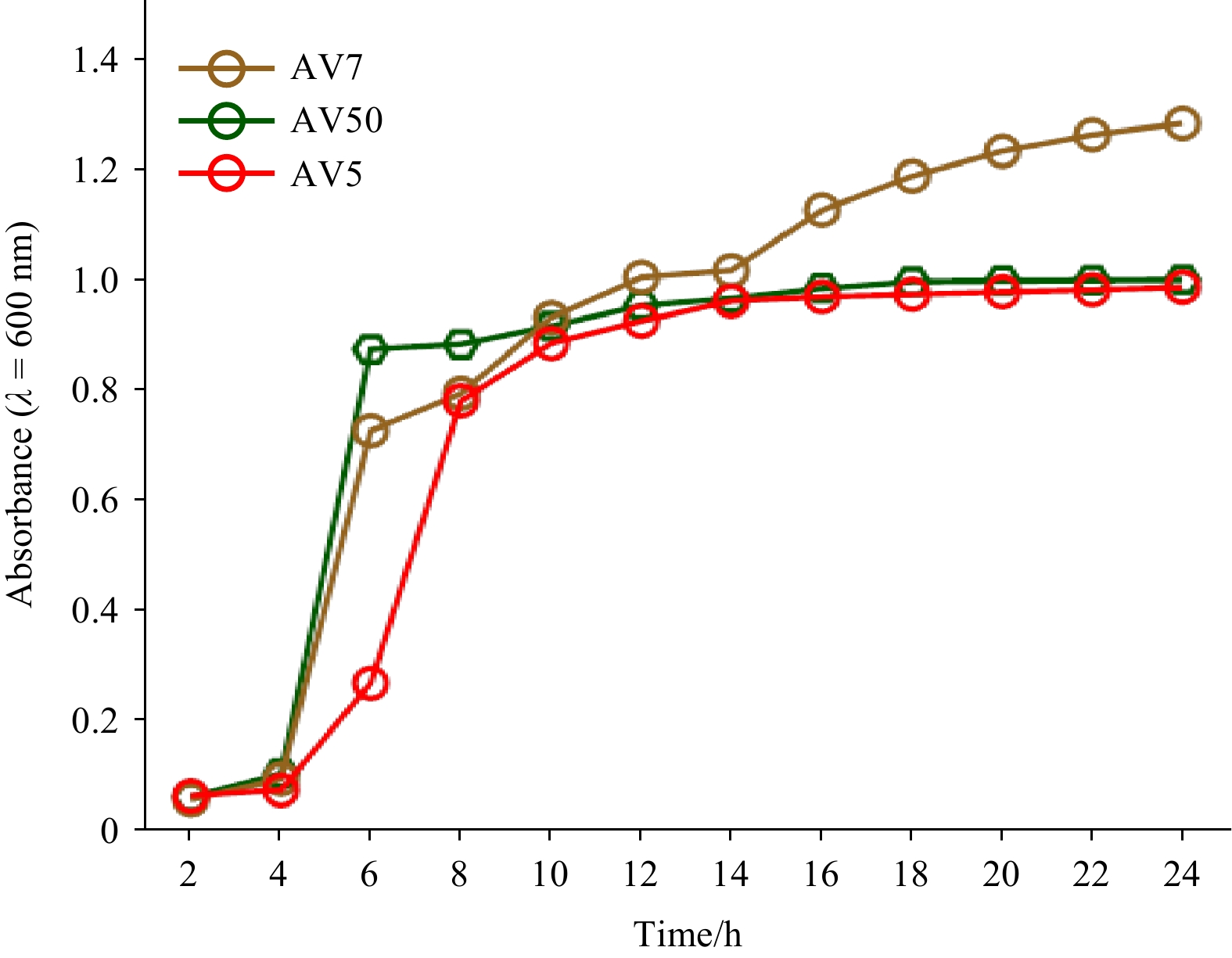
We observed the effects of temperature and agitation on the growth of the bacteria at 37℃ and 150 r/min. As shown in Fig. 2, the three isolated strains (AV5, AV7, and AV50) exhibited different growth patterns. After 2 h, all three isolates entered the stationary phase at 37℃ and 150 r/min, which indicates that they had reached the maximum growth rate. However, AV50 and AV5 reached the stationary phase earlier than AV7 (Fig. 2).
Figure
2.
The growth phases of the selected strains (B. amyloliquefaciens AV5, B. subtilis AV7, and B. velezensis AV50) determined at an absorbance wavelength of 600 nm after incubation for 24 h.
3.3
Morphological and biochemical features of probiotic isolates
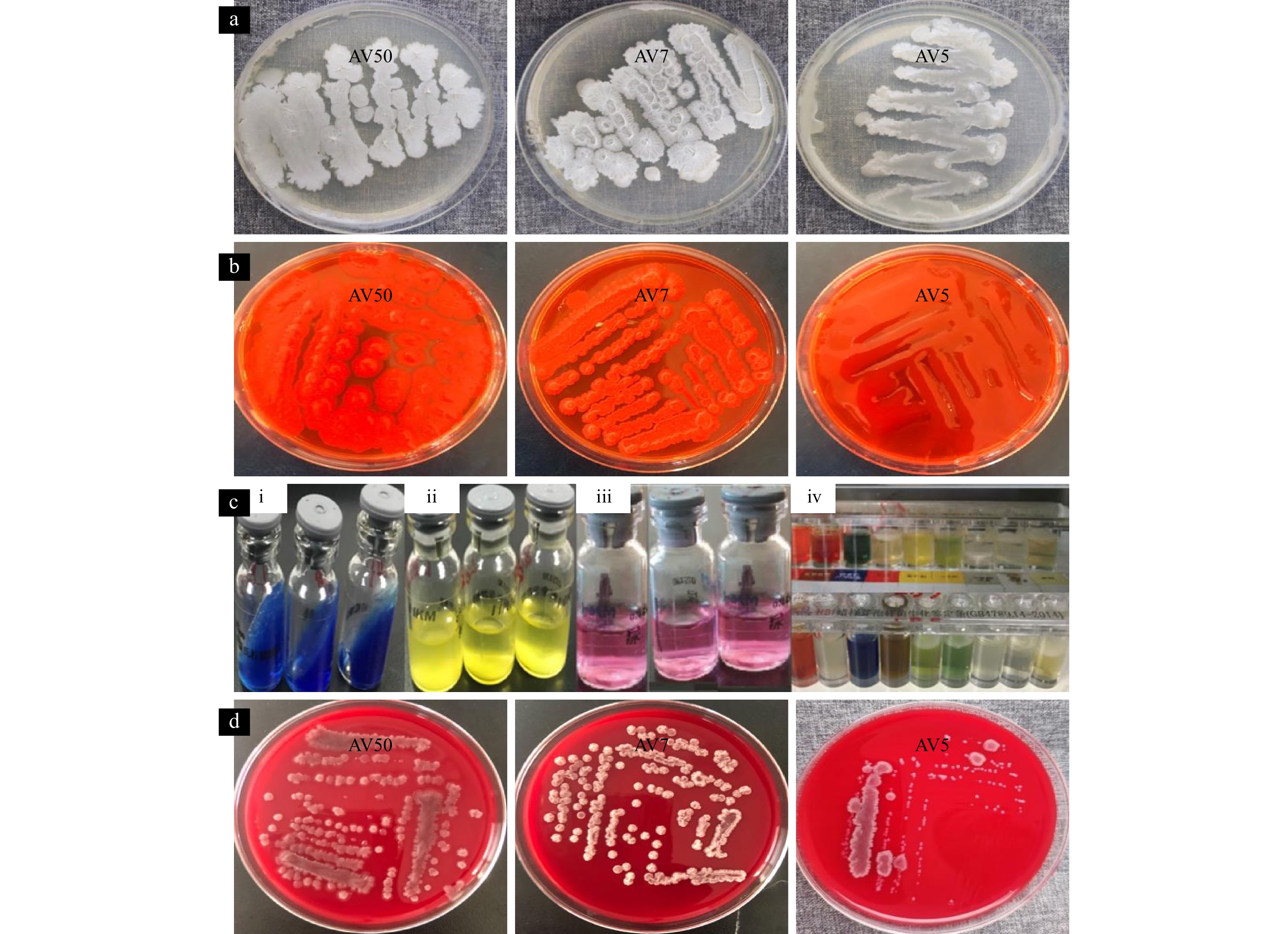
Tables 1 and 2 illustrate that the three Bacillus isolates possessed unique morphological and biochemical characteristics. Despite their morphological differences, all three isolates shared a significant number of biochemical properties, notably their ability to utilize almost all tested carbon sources. However, AV50 does not utilize certain carbon sources, such as citrate. None of the isolates demonstrated the ability to ferment lactose, produce hippuric acid, liquefy gelatin, produce methyl red, or show Voges-Proskauer (VP) activity. Among the three, only AV7 could grow in lysozyme broth and tested positive for mannitol utilization, indicating its halophilic nature (Fig. 3).
Figure
3.
Morphological, biochemical, and hemolytic properties of the selected isolates. a. Morphology of AV50, AV7, AV5; b. biofilm formation of isolates (AV7, AV50, and AV50) using the Congo red agar method; c. evidence of biochemical test of the isolates (i: citrate reduction test, ii: urease test, iii: arginine dihydrolase test, and iv: a confirmatory test using Bacillus cereus identification bar); and d. hemolytic activity of AV50, AV7, and AV5.
The isolated Bacillus strains were evaluated for safety to ensure their suitability as probiotics. The haemolytic activities of AV5, AV7, and AV50 were further analyzed to determine the degree of haemolysis, revealing that all three compounds displayed γ-haemolysis, with AV5 showing the most significant activity in the present study (Fig. 3).
3.5
Biosafety assay
In the in vitro biosafety assessments, neither the experimental nor the control fish displayed any pathological symptoms (edema, hemorrhage, lesions, scaly loss, or mucus). No mortality was observed, it is important to note that the absence of mortality in the study suggests that the isolates were not harmful to the organisms being tested, therefore isolates were not pathogenic.
3.6
Antibiotic susceptibility
Table 3 presents the antibiotic susceptibility results for the selected isolates. A total of 20 antibiotics were tested, and all isolates were highly susceptible (S) to 17 antibiotics. The AV7 strain demonstrated resistance (R) to furazolidone. In contrast, strains AV50 and V5 showed MS only for sulfamethoxazole and polymyxin b, respectively.
3.7
Bile salt tolerance
The percentage of survivors after three hours of exposure was greater than 50%. However, after 4 h of bile salt treatment, the durability rate of AV50 declined to 47.7%. Simultaneously, the survivability rates of AV7 and AV5 continued to exceed 50%. (Fig. 4). AV50 exhibited a significant (p<0.05) decline in viability every hour after 2 h of exposure to bile salts. After 1 h and 2 h, AV7 showed a significant decline in percentage survival (94.3% and 73.2%, respectively), but the decrease persisted until the 4th hour. In AV5 cells, a significant decline in cell viability was observed after 2 h, which remained consistent thereafter.
Figure
4.
The findings are shown as the mean±SD (n=3) of the isolates tested for bile tolerance (0.5%) at 37℃ after 4 h. Based on Tukey’s test, values with distinct superscripts on the same line show a significant difference (p˂0.05). AV5 represents B. amyloliquefaciens, AV7 B. subtilis, and AV50 B. velezensis.
Three isolates showed promising results after exposure to 10 min, 5 min, and 2 min for 100℃, 90℃, and 80℃, respectively. Compared to the control, all isolates subjected to various temperatures displayed high growth (CFU). As shown in Fig. 5, a significant difference (p<0.05) was observed at the different exposure time points. CFU values increased significantly (p<0.05) when AV5 was subjected to 80℃ for 2 min versus 90℃ and 100℃, 100℃ for 5 min versus 80℃ and 90℃, and 90℃ and 100℃ for 10 min versus 80℃. The AV50 strain exhibited significantly higher (p<0.05) CFU values when heated at 80℃ and 90℃ for 2 min versus 100℃, 90℃ for 5 min versus 80℃ and 100℃, and 90℃ for 10 min versus 80℃ and 100℃. In contrast, strain AV7 exhibited significantly higher CFU values (p<0.05) when subjected to 90℃ for 2 min versus 80℃ and 100℃, 100℃ for 5 min versus 80℃ and 90℃, and 80℃ for 10 min versus 90℃ and 100℃.
Figure
5.
The findings are shown as the mean±SD (n=3) of the isolates tested for temperature resistance (80℃, 90℃, and 100℃). Based on Tukey's test, values with distinct superscripts on the same line show a significant difference (p˂0.05). AV5 represents B. amyloliquefaciens, AV7 B. subtilis, and AV50 B. velezensis.
No suppression was observed among the three strains during the compatibility experimental tests, indicating that the three isolates were sufficiently compatible to be combined (multispecies characteristics) to exhibit their beneficial effects without causing any detrimental effects on the host organism.
3.10
Antimicrobial test
All three isolates inhibited at least one of the pathogens tested (Table 4). The AV50 strain showed no inhibition of S.iniae (SI) or V. harveyi (VH) in both cross-streak and agar well diffusion assays. However, the AV50 strain exhibited 3–4 mm inhibition of V. alginolyticus (VA) and S. agalactiae (SA). In comparison, strains AV7 and AV5 demonstrated higher antimicrobial activity, with inhibition zones ranging from 5 mm to 6 mm. Notably, AV5 exhibited the highest antimicrobial activity with inhibition zones exceeding 7 mm (Fig. 6).
Table
4.
Antimicrobial assay of selected Bacillus strains against pathogenic bacteria
Pathogenic isolate
Inhibition zone of isolates/mm
AV5
AV7
AV50
Vibrio harveyi
7.3 ± 0.3a
6.981 ± 0.2a
4.98 ± 0.2b
Vibrio alginolyticus
3.87 ± 0.2a
6.2 ± 0.1b
2.3 ± 0.1c
Streptococcus agalactiae
7.5 ± 0.1a
7.3 ± 0.1a
3.4 ± 0.2b
Streptococcus iniae
3.6 ± 0.1a
2.8 ± 0.1b
1.4 ± 0.1b
Note: Results are presented as the mean ± SD (n=3). Based on Tukey's test, values with distinct superscripts on the same line show a significant difference (p˂0.05). The inhibition zone was determined by subtracting the well diameter (mm) from the inhibition zone diameter (mm). Thus, the following variables are defined: B. amyloliquefaciens (AV5), B. velezensis (AV7), and B. velezensis (AV50).
Figure
6.
An illustration displaying the inhibition zones of the selected strains (AV7, AV50, and AV5) against the fish pathogens (Vibrio harveyi, VH; Vibrio alginolyticus, VA; Streptococcus agalactiae, SA; and Streptococcus iniae, SI) using the agar well-diffusion method in Lane 1 (L1, a-l) and cross streak method in Lane 2 (L2). AV5 represents B. amyloliquefaciens, AV7 B. subtilis, and AV50 B. velezensis.
3.11
Hydrophobicity properties of the isolated strains
The adhesion of the selected isolates (AV5, AV7, and AV50) was measured using ethyl acetate, chloroform, and xylene (Fig. 7a). Compared to xylene and chloroform, isolates AV5 and AV7 displayed significantly lower hydrophobicity (p<0.05) to ethyl acetate.
Figure
7.
The findings are shown as the mean± SD (n=3). a. The hydrophobicity (%) of the cell surfaces of the selected isolates versus solvents (ethyl acetate, chloroform, and xylene), and b. auto-aggregation tests of the three strains after (24 h). Based on Tukey's test, values with distinct superscripts on the same line show a significant difference (p˂0.05). AV5 represents B. amyloliquefaciens, AV7 B. subtilis, and AV50 B. velezensis.
In the digestive tract, auto-aggregation was highly correlated with cell adhesion, and all three isolates, AV5, AV7, and AV50, showed poor adhesion after 3 h (less than 30%). In contrast, after 24 h, the cell adhesion was 92.66%, 84.67%, and 89.33%, respectively (Fig. 7b). It is important to note that these results suggest that cell adhesion increased significantly after 24 h, with the most notable improvement observed in the group with the highest initial adhesion rate, which increased to 92.66%, indicating that auto-aggregation was strongly associated with cell adhesion in the digestive tract.
3.13
Growth and pH test of the isolated strains
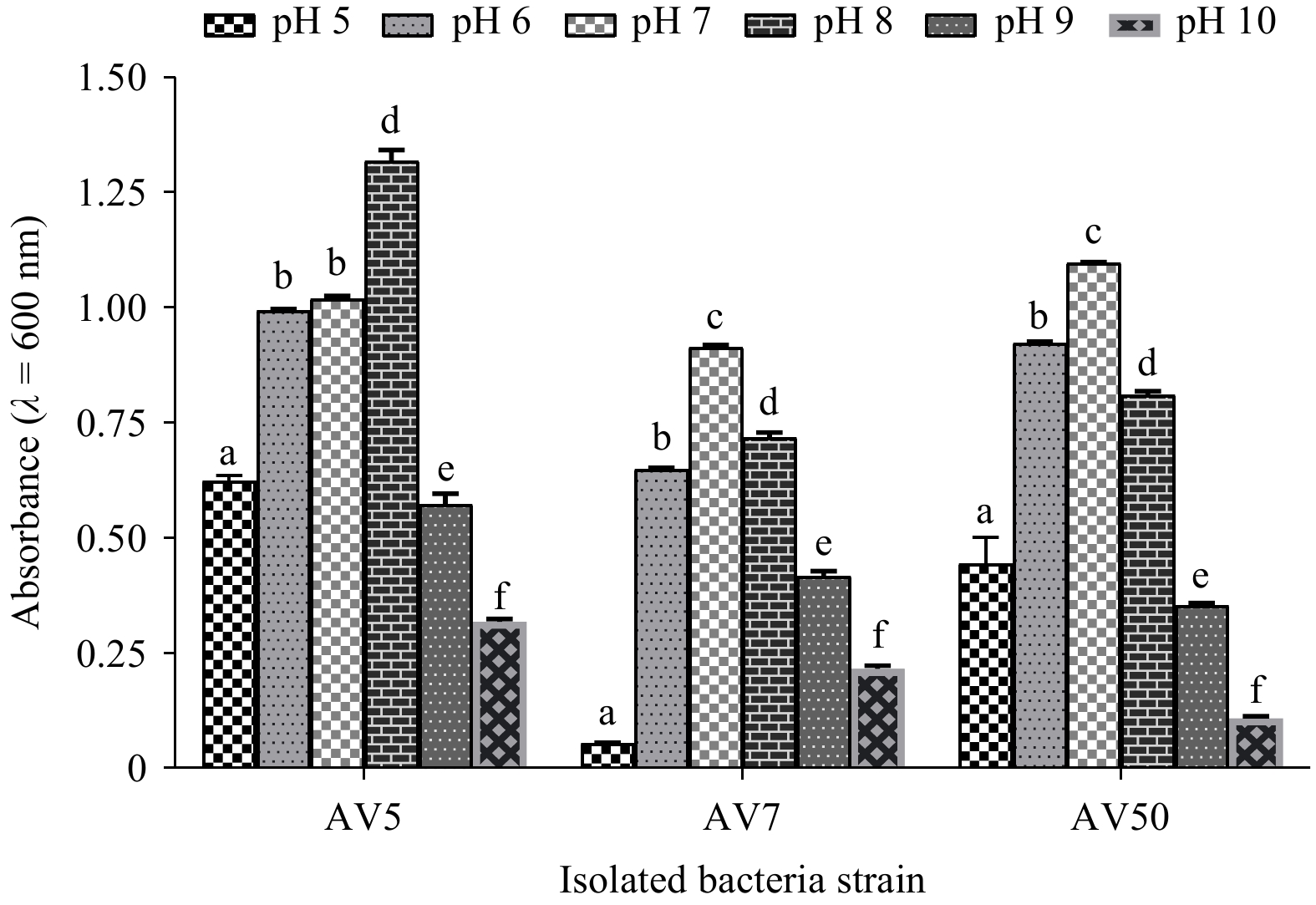
Isolates AV5, AV7, and V50 exhibited promising tolerance. However, vegetative cell growth gradually increased within the pH range of 1.0–8.0, with the AV5 and AV50 strains showing optimum growth between pH 5.0 and 8.0, and the AV7 strain showing optimum growth between pH 6.0 and 8.0. No significant difference (p>0.05) in the vegetative cell growth of strain AV5 was observed between pH 5.0 and 8.0. As the pH increased from 9.0 to 10.0, growth decreased, indicating that the isolated strains survived under acidic and alkaline conditions. The isolated strains showed significant differences (p<0.05) in growth at different pH levels (Fig. 8).
Figure
8.
The findings are shown as the mean±SD (n=3) of the selected isolates’ viability at pH (5.0–10.0). Based on Tukey’s test, values with distinct superscripts on the same line show a significant difference (p˂0.05). AV5 represents B. amyloliquefaciens, AV7 B. subtilis, and AV50 B. velezensis.
Biofilm formation was screened and analyzed using the Congo red method. In the current study, none of the isolates exhibited biofilm formation after the test, as evidenced by the absence of black colonies (Table 2 and Fig. 3), indicating that the isolated strains had a strong ability to adhere, survive, and compete in the gastrointestinal tract.
4.
Discussion
Efficient disease management in aquaculture is crucial for the sustainable and successful production of aquatic animals (Abarike et al., 2018). The widespread and improper use of antibiotics has raised significant biological and ecological concerns, particularly regarding the emergence of antibiotic-resistant bacteria (Yang et al., 2019). Beneficial microbes, known as probiotics, are recommended as antibiotic substitutes because they promote health, effectiveness, environmental friendliness, and pathogen resistance (Vijayaram et al., 2024). Probiotic Bacillus sp. exhibits various characteristics, including the production of non-pathogenic substances, enhancement of water quality, heat tolerance, extended shelf life, and resilience under harsh conditions owing to their sporulation capability (Kuebutornye et al., 2019; Amoah et al., 2021). In aquaculture, Probiotic Bacillus sp. are isolated from diverse sources, such as decaying matter, water, commercial sources, fish and vertebrate gastrointestinal tracts, and soil. Recently, Bacillus sp. have shown beneficial effects in aquaculture, with multiple studies highlighting their positive effects on Nile tilapia cultivation (Shija et al., 2023). Researchers typically choose Bacillus spp. among the many probiotics employed because of their ability to remain alive for a long time and their resilience to UV radiation, severe temperatures, and pH. Several studies have been published on the benefits of Bacillus spp. in aquaculture animals (Kuebutornye et al., 2019; Shija et al., 2023). According to previous reports, Bacillus probiotics related to hosts, particularly those found in the intestines of healthy fish, can survive and establish themselves in the host more efficiently. In addition, they are capable of providing health benefits (Van Doan et al., 2018; Ramesh et al., 2015). However, few studies have focused on screening and characterizing probiotics derived from the gastrointestinal tract of freshwater animals, particularly the Nile tilapia. Therefore, in this study, the probiotic potential of three Bacillus sp. (B. amyloliquefaciens AV5(OR647358), B. subtilis AV7 (LC781790), and B. velezensis AV50 (OR647359)) isolated from Nile tilapia intestines was investigated. The strains AV5, AV7, and AV50 were identified based on morphological traits and biochemical tests and confirmed by 16S rDNA gene sequencing. All isolates demonstrated the ability to utilize various carbon sources, including amino acids, such as arginine, inositol, adonitol, citrate, gelatin, lactose, starch, rhamnose, sorbitol, and glucose. These characteristics suggest their potential utility as probiotics and in the production of value-added products in the food industry, as documented in previous studies (Lee et al., 2017; Kavitha et al., 2018; Kuebutornye et al., 2020). In contrast to other probiotics, Bacillus strains produce heat-resistant spores (Kuebutornye et al., 2020; Amoah et al., 2021), and are resistant to low pH and high bile concentrations (Šimunović et al., 2022). For a probiotic bacterium to be considered as potentially fulfilling the biological function of promoting animal and human health, it must be able to survive and remain viable despite encountering challenging environmental conditions, such as high levels of bile, low pH in the stomach, and high temperatures during passage through the gastrointestinal tract (Amenyogbe et al., 2024). They have also demonstrated the ability to thrive in the gastrointestinal tract of fish (Šimunović et al., 2022). According to García-Ruiz et al. (2014), there is a high survival rate of Bacillus sp. in the human gut with varying bile contents (0.3%–0.5%). The stomach is essential for food digestion, releasing gastric juices composed of mucus, hydrochloric acid (HCl), water, pepsin, and intrinsic factors. Pepsin, the main enzyme, hydrolyzes proteins into smaller peptides and amino acids but requires the acidic environment provided by HCl to function effectively within a pH range of 1.0 to 3.0. Any probiotic must withstand this specific pH range (Ergun et al., 2022; Bardhan et al., 2012). The ability to withstand harsh heat conditions, often used in animal feed production for pathogen elimination and flavor enhancement, is a crucial biotechnological trait of probiotics (Guo et al., 2016). In our study, all isolated strains (AV5, AV7, and AV50) demonstrated high sporulation efficiency, enabling them to survive at 0.5% bile concentration (Fig. 5), pH as low as 1 (Fig. 8), and post-heat treatment, surpassing the control group (Fig. 6). Furthermore, higher heat treatments (80℃ to 100℃) activated the selected isolates, promoting faster growth. An earlier study posited that Bacillus sp. exposed to temperatures of 80℃, 90℃, and 100℃ exhibited significant bacterial growth (Kort et al., 2005). This observation is consistent with the findings of the current study on the thermal tolerance of Bacillus isolates and previous research on the sporulation capabilities of Bacillus sp. including B. velezensis, B. stercoris, and B. subtilis (Amoah et al., 2024). Probiotics are widely used in the food and feed sectors because of their health benefits. The findings of this study suggest that the newly isolated strains possess notable biotechnological potential owing to their ability to endure extreme heat. Commercial animal feed manufacturing relies on heat to achieve high digestibility and eliminate pathogens. Therefore, being thermotolerant is a positive sign that it may be used as feed in the aquaculture industry.
When selecting a probiotic, its ability to adhere to the intestinal surface and epithelial cells should be considered. This ensures that they can withstand intestinal changes and inhibit harmful bacteria by occupying intestinal spaces (Liu et al., 2017). Consequently, probiotics may restore the microbial balance in the gut, increase beneficial bacteria (Collado et al., 2007), and enhance the host's immune system (Nikoskelainen et al., 2001).
Bacillus velezensis (Ye et al., 2018), B. subtilis (Guo et al., 2016), and B. amyloliquefaciens (Kuebutornye et al., 2020) have been extensively studied for their sporulation ability. Bacillus sp. possesses a crucial ability to colonize and adhere to epithelial cells and mucosal surfaces, thereby ensuring resistance to alterations in intestinal contents and impeding the attachment of harmful bacteria throughout the intestine, ultimately minimizing inflammation (Shinde et al., 2019; Rohith Halami, 2021; Li et al., 2020).
Hydrophobicity helps to analyze its ability to adhere to the host's intestinal epithelial lining (Kuebutornye et al., 2020). In the current study, AV5 exhibited significantly higher hydrophobicity (97.5% and 97.1% with chloroform and xylene, respectively) than AV50 (92.8% and 95.8%) and AV7 (85.1% and 96.6%, respectively), indicating superior attachment to hydrocarbons. Also, AV7 showed a higher hydrophobicity than AV50 and AV5 with ethyl acetate (90.6%). Our study found significantly higher hydrophobicity results compared to those of Lee et al. (2017), suggesting enhanced electron donation and acceptance for better epithelial cell adhesion. In addition, auto-aggregation activity is directly proportional to cell adhesion in the digestive tract, which is a prerequisite for beneficial probiotic bacteria (Kuebutornye et al., 2020).
Bacterial strains that cause aggregation result in the formation of a precipitate, leading to a clear upper solution, whereas non-aggregating bacteria result in a turbid suspension (Balakrishna and Kumar, 2012). In the present study, AV5, AV7, and AV50 exhibited significant auto-aggregation rates of 92.66%, 84.67%, and 89.33%, respectively, consistent with previous studies (Kuebutornye et al., 2020). Furthermore, there is a significant correlation between auto-aggregation and the adherence of cells to the digestive system, which is an essential quality for an effective probiotic (Liu et al., 2017). This study revealed that the isolated strains AV5, AV7, and AV50 exhibited significant auto-aggregation, which aligns with previous studies (Wang et al., 2021; Dunlap et al., 2020).
Another vital aspect for identifying potential probiotic bacteria in vitro is the evaluation of hemolytic activity, compatibility, and biofilm formation (Kuebutornye et al., 2020).
The presence or absence of haemolytic activity is one of the most significant safety pre-conditions during the selection and characterization of a potential probiotic strain. Haemolysis is used to describe the breakdown of red blood cells. Hemolysin, one of the best-categorized virulence factors, is toxic to endothelial, epithelial, and phagocytic cells (Gottschalk Segura, 2000). When produced by pathogenic bacteria, hemolysin is reported to induce the lysis of host cells, which later leads to the release of iron-containing compounds (including haemoglobin) that are valuable for the host organism’s bacterial growth (Argyri et al., 2013). Either γ-hemolysis or β-hemolysis is safe, whereas α-hemolysis is harmful (Deng et al., 2023). In the current study, all strains exhibited γ-hemolysis, indicating non-hemolytic behavior and enhancing the safety of host organisms, which aligns with the results of previous studies (Lee et al., 2017; Kavitha et al., 2018).
Compatibility testing is mostly undertaken to determine whether the isolated strains are suitable for use as multi-species probiotics (Kuebutornye et al., 2020). Compatibility testing helps ascertain whether the isolated strains may be merged and used as multispecies probiotics. A previous study conducted by Saarela et al. (2000), showed that food products made from probiotics consisting of a single species had an acidic and sour taste, unlike those made from many species (Saarela et al., 2000). In the current study, all three isolates showed mutual compatibility among strains AV5, AV7, and AV50, indicating their ability to be used as multispecies (Amoah et al., 2024). Also, Kuebutornye et al. (2020) similarly observed comparable compatibility outcomes across Bacillus species.
On the other hand, despite the numerous advantages of using biofilm-producing bacteria like Bacillus subtilis in public health, there have been indications that these bacteria are less susceptible to antimicrobial agents and play a more significant role in causing human diseases. This highlights the need for conducting a biofilm formation test (Kuebutornye et al., 2020). The present study revealed that all three isolates (AV5, AV7, and AV50) yielded negative results in the biofilm-producing assays, which is consistent with prior studies (Kuebutornye et al., 2020; Kavitha et al., 2018). However, out of the three isolates examined in the earlier studies conducted by Kavitha et al. (2018), only one strain demonstrated a positive result for biofilm formation.
To be labeled as a probiotic, a bacterium must be non-harmful to the host organism and incapable of transmitting antibiotic-resistance genes (Elshaghabee et al., 2017; Ouwehand et al., 2016). In addition to the previously described in vitro safety experiments, we also performed biosafety assessment tests using the in vivo challenge trial. The biosafety test revealed that all three isolates (AV5, AV7, and AV50) were found to be non-pathogenic, indicating that they are safe for Nile tilapia, and the results were consistent with previous studies of Kuebutornye et al. (2020). Also, the lack of disease-causing ability of Bacillus subtilis (Kuebutornye et al., 2020), B. stercoris (Wang et al., 2021), and B. velezensis (Gao et al., 2017) has been shown.
Moreover, a prospective probiotic bacterium must not possess acquired and transferable antibiotic-resistance genes (Cabello, 2006). The European Food Safety Authority (EFSA) has found that the two antibiotic groups most commonly recommended for in vitro testing to select beneficial strains are those that hinder cell wall synthesis (ampicillin and vancomycin) and those that hinder protein synthesis (gentamycin, tetracycline, chloramphenicol, streptomycin, erythromycin, clindamycin, and kanamycin) (EFSA Panel on Additives and Products or Substances used in Animal Feed, 2012), which were also tested in the present study. Also, the EFSA has recommended that, if feasible, the focus for selecting bacteria as feed additives should be on those that are less resistant.
Out of the 20 antibiotics tested, the three isolates were found to be highly susceptible to 17 of them. These antibiotics include tetracycline, erythromycin, chloramphenicol, penicillin, ciprofloxacin, kanamycin, ampicillin, gentamicin, clindamycin, ceftriaxone, cefoperazone, doxycycline, vancomycin, midecamycin, ofloxacin, piperacillin, and norfloxacin, highlighting their potential safety and efficacy in aquaculture settings (Guo et al., 2016; Kuebutornye et al., 2020). However, only B. velezensis strain AV50 showed resistance to furazolidone antibiotic. Moderate susceptibility was observed in polymyxin b antibiotic for B. velezensis AV50 and B. amyloliquefaciens AV5 strains, and sulfamethoxazole antibiotic for only strain B. subtilis AV7.
Notably, AV7 exhibited resistance to these antibiotics possibly because of the presence of natural resistance genes (Larsen et al., 2014). There are several viewpoints suggesting that some strains of probiotics can withstand particular medications and live when taken with antibiotic treatment (Tarale et al., 2015). However, there has been a growing interest in the connection between the use of antimicrobials in animal feeding and the emergence of antibiotic-resistant microbes in humans (Salyers et al., 2004). The current investigation corroborates with the study conducted by Kuebutornye et al. (2020), who reported that B. velezensis isolated from the gut of hybrid grouper exhibited resistance to the furazolidone antibiotic. In addition, B. amyloliquefaciens obtained from the intestinal tract of tilapia (O. niloticus) (Kuebutornye et al., 2020), showed a moderate susceptibility to furazolidone antibiotics.
Another compelling health benefit of probiotics is their capacity to prevent diseases. In the present study, the isolated strains AV5, AV7, and AV50 also demonstrated significant antagonistic effects against common fish pathogens, such as Vibrio alginolyticus, Streptococcus iniae, V. harveyi, and S. agalactiae, suggesting their potential for disease prevention and enhancing aquaculture sustainability, which aligns with previous results (Bluford et al., 2017; Shoemaker et al., 2001; Bowater et al., 2012). The results suggest that the three strains AV5, AV7, and AV50 possess the ability to combat fish diseases, thereby promoting the sustainability of the aquaculture sector.
Several infections that are known to hinder fish productivity have been attributed to Vibrio spp. (Bluford et al., 2017) and Streptococcus spp. (Shoemaker et al., 2001). For example, reports indicated that a severe infection caused by S. agalactiae was observed in giant groupers in Australia in 2012 (Bowater et al., 2012).
The results of this investigation demonstrated that the examined strains exhibited distinct zones of inhibition to different extents when cultured in pathogenic broth, indicating that the three isolates were capable of restricting the development of pathogenic bacteria. Previous works have provided evidence that Bacillus species have superior antibacterial capabilities against pathogenic microorganisms. One example is B. velezensis CC09, which has a broad range of antibacterial activity and has great promise for use in biocontrol and biotechnology. This activity is mainly due to the production of polyketides, lipopeptides, and antibacterial proteins (Cai et al., 2017). Lipopeptides have garnered significant interest owing to their several advantageous characteristics, including low toxicity, potent antibacterial and antiviral activities, excellent endurance to high temperatures, and exceptional biodegradation capability (Zhao et al., 2014). These results indicate that the three Bacillus strains AV5, AV7, and AV50 can potentially combat fish diseases.
5.
Conclusions
This work highlights the impressive probiotic capabilities of three Bacillus strains, AV5, AV7, and AV50, isolated from the intestine of Nile tilapia (O. niloticus). The isolates exhibited remarkable characteristics, including strong resilience under simulated gastric conditions that replicated the digestive environment of humans, ensuring their capacity to survive and operate well in the gastrointestinal tract. Moreover, their displayed characteristics, such as their ability to survive high temperatures, non-haemolytic nature, broad antimicrobial activity, and susceptibility to antibiotics, make them promising candidates for probiotics. The utilization capabilities of isolates for different carbon sources, such as gelatin, nitrate, arginine amino acids, lactose, glucose, starch, adonitol, rhamnose, sorbitol, and inositol, may potentially assist in the breakdown and digestion of carbohydrates and amino acids. Considering these findings, AV5, AV7, and AV50 show significant promise as probiotics in aquaculture. However, to ensure suitability for use in Nile tilapia aquaculture, additional in vitro and in vivo trials are necessary to verify the efficacy of the proposed methods.
Acknowledgments:
The authors express our gratitude to all the laboratory members for their positive criticisms and suggestions, making this study genuine.
Ze Meng, Lei Zhou, Baosheng Li, Jianhuang Qin, Juncheng Xie. Erratum to: Acta Oceanologica Sinica (2022) 41(10): 119–130DOI: 10.1007/s13131-022-2023-3The atmospheric hinder for intraseasonal sea-air interaction over the Bay of Bengal during Indian summer monsoon in CMIP6[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-022-2131-0
Ze Meng, Lei Zhou, Baosheng Li, Jianhuang Qin, Juncheng Xie. Erratum to: Acta Oceanologica Sinica (2022) 41(10): 119–130DOI: 10.1007/s13131-022-2023-3The atmospheric hinder for intraseasonal sea-air interaction over the Bay of Bengal during Indian summer monsoon in CMIP6[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-022-2131-0
Table
1.
Morphological features of the isolates from the intestine of the Nile tilapia (Oreochromis niloticus)
Isolate
Color
Form
Surface
Size
Margin
Elevation
Texture
AV7
white
circular
dry
medium
entire
raised
rough
AV5
creamy white
irregular
moist
medium
undulate
umbonate
rough
AV50
white
irregular
dry
medium
undulate
umbonate
rough
Note: The three isolated strains, AV5, AV7, and AV50, exhibited identity sequence homology (99%) to B. amyloliquefaciens, B. subtilis, and B. velezensis respectively.
Table
2.
The biochemical features of the selected strains isolated from the intestine of the Nile tilapia (Oreochromis niloticus)
Tests performed
Biochemical feature
Tests performed
Biochemical feature
AV50
AV7
AV5
AV50
AV7
AV5
Mannitol
+
−
−
Simon’s citrate
+
−
+
Lactose fermentation
–
–
–
Starch hydrolysis
+
+
+
Gram staining
+
+
+
Biofilm production
–
–
–
Glucose
+
+
+
Spore formation
+
+
+
Rhamnose
+
+
+
Arginine dihydrolase
+
+
+
Nitrate reduction
+
+
+
Sorbitol
+
+
+
Lysozyme broth
–
+
–
Inositol
+
+
+
Methyl red
–
–
–
Adonitol
+
+
+
Urease
+
+
+
Haemolysis
$\gamma $
$\gamma $
$\gamma $
Gelatin liquefaction
–
–
–
Catalase
+
+
+
Hippuric acid
–
–
–
Vorges proskauer
–
–
–
Note: The three isolated strains, AV5, AV7, and AV50, exhibited identity sequence homology (99%) to B. amyloliquefaciens, B. subtilis, and B. velezensis respectively. Additionally, + donated a positive response, – a negative response, and γ no response.
Table
3.
Sensitivity of isolates (mm) to antibiotics
Antibiotic
Antibiotic content/ µg·disc-1
Isolate
AV5
AV7
AV50
Tetracycline
30
S
S
S
Erythromycin
15
S
S
S
Chloramphenicol
30
S
S
S
Polymyxin B
300
S
S
MS
Penicillin
10
S
S
S
Ciprofloxacin
5
S
S
S
Kanamycin
30
S
S
S
Ampicillin
10
S
S
S
Gentamicin
10
S
S
S
Furazolidone
300
S
R
S
Clindamycin
2
S
S
S
Ceftriaxone
30
S
S
S
Cefoperazone
75
S
S
S
Doxycycline
30
S
S
S
Vancomycin
30
S
S
S
Midecamycin
30
S
S
S
Ofloxacin
5
S
S
S
Sulfamethoxazole
27
MS
S
S
Piperacillin
100
S
S
S
Norfloxacin
10
S
S
S
Note: Isolate antibiotics sensitivity was evaluated according to the McFarland standard, incubated at 37℃ for 24 h, and the susceptibility of the isolates was determined by measuring the diameter of the inhibition zone (in millimeters). S, R, and MS donate susceptible, resistant, and moderately susceptible responses respectively. B. amyloliquefaciens (AV5), B. velezensis (AV7), and B. velezensis (AV50).
Table
4.
Antimicrobial assay of selected Bacillus strains against pathogenic bacteria
Pathogenic isolate
Inhibition zone of isolates/mm
AV5
AV7
AV50
Vibrio harveyi
7.3 ± 0.3a
6.981 ± 0.2a
4.98 ± 0.2b
Vibrio alginolyticus
3.87 ± 0.2a
6.2 ± 0.1b
2.3 ± 0.1c
Streptococcus agalactiae
7.5 ± 0.1a
7.3 ± 0.1a
3.4 ± 0.2b
Streptococcus iniae
3.6 ± 0.1a
2.8 ± 0.1b
1.4 ± 0.1b
Note: Results are presented as the mean ± SD (n=3). Based on Tukey's test, values with distinct superscripts on the same line show a significant difference (p˂0.05). The inhibition zone was determined by subtracting the well diameter (mm) from the inhibition zone diameter (mm). Thus, the following variables are defined: B. amyloliquefaciens (AV5), B. velezensis (AV7), and B. velezensis (AV50).
Figure 1. A phylogenetic tree based on 16SrDNA sequences generated by the neighbor-joining method (1 000 bootstraps) using the NCBI distance tree. This figure illustrates the relationship between closely related Bacillus species and selected Bacillus strains (AV5, AV7, and AV50).
Figure 2. The growth phases of the selected strains (B. amyloliquefaciens AV5, B. subtilis AV7, and B. velezensis AV50) determined at an absorbance wavelength of 600 nm after incubation for 24 h.
Figure 3. Morphological, biochemical, and hemolytic properties of the selected isolates. a. Morphology of AV50, AV7, AV5; b. biofilm formation of isolates (AV7, AV50, and AV50) using the Congo red agar method; c. evidence of biochemical test of the isolates (i: citrate reduction test, ii: urease test, iii: arginine dihydrolase test, and iv: a confirmatory test using Bacillus cereus identification bar); and d. hemolytic activity of AV50, AV7, and AV5.
Figure 4. The findings are shown as the mean±SD (n=3) of the isolates tested for bile tolerance (0.5%) at 37℃ after 4 h. Based on Tukey’s test, values with distinct superscripts on the same line show a significant difference (p˂0.05). AV5 represents B. amyloliquefaciens, AV7 B. subtilis, and AV50 B. velezensis.
Figure 5. The findings are shown as the mean±SD (n=3) of the isolates tested for temperature resistance (80℃, 90℃, and 100℃). Based on Tukey's test, values with distinct superscripts on the same line show a significant difference (p˂0.05). AV5 represents B. amyloliquefaciens, AV7 B. subtilis, and AV50 B. velezensis.
Figure 6. An illustration displaying the inhibition zones of the selected strains (AV7, AV50, and AV5) against the fish pathogens (Vibrio harveyi, VH; Vibrio alginolyticus, VA; Streptococcus agalactiae, SA; and Streptococcus iniae, SI) using the agar well-diffusion method in Lane 1 (L1, a-l) and cross streak method in Lane 2 (L2). AV5 represents B. amyloliquefaciens, AV7 B. subtilis, and AV50 B. velezensis.
Figure 7. The findings are shown as the mean± SD (n=3). a. The hydrophobicity (%) of the cell surfaces of the selected isolates versus solvents (ethyl acetate, chloroform, and xylene), and b. auto-aggregation tests of the three strains after (24 h). Based on Tukey's test, values with distinct superscripts on the same line show a significant difference (p˂0.05). AV5 represents B. amyloliquefaciens, AV7 B. subtilis, and AV50 B. velezensis.
Figure 8. The findings are shown as the mean±SD (n=3) of the selected isolates’ viability at pH (5.0–10.0). Based on Tukey’s test, values with distinct superscripts on the same line show a significant difference (p˂0.05). AV5 represents B. amyloliquefaciens, AV7 B. subtilis, and AV50 B. velezensis.



 DownLoad:
DownLoad:
 DownLoad:
DownLoad:









 DownLoad:
DownLoad: