Temim Deli, Noureddine Chatti, Khaled Said, Enrique González-Ortegón, Christoph D. Schubart. Population genetic polymorphism and structure in the two coastal crab species Carcinus aestuarii (Brachyura, Carcinidae) and Pachygrapsus marmoratus (Brachyura, Grapsidae), across the Mediterranean Sea, reflect residual effects of different evolutionary histories[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-024-2401-1
Citation:
Temim Deli, Noureddine Chatti, Khaled Said, Enrique González-Ortegón, Christoph D. Schubart. Population genetic polymorphism and structure in the two coastal crab species Carcinus aestuarii (Brachyura, Carcinidae) and Pachygrapsus marmoratus (Brachyura, Grapsidae), across the Mediterranean Sea, reflect residual effects of different evolutionary histories[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-024-2401-1
Temim Deli, Noureddine Chatti, Khaled Said, Enrique González-Ortegón, Christoph D. Schubart. Population genetic polymorphism and structure in the two coastal crab species Carcinus aestuarii (Brachyura, Carcinidae) and Pachygrapsus marmoratus (Brachyura, Grapsidae), across the Mediterranean Sea, reflect residual effects of different evolutionary histories[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-024-2401-1
Citation:
Temim Deli, Noureddine Chatti, Khaled Said, Enrique González-Ortegón, Christoph D. Schubart. Population genetic polymorphism and structure in the two coastal crab species Carcinus aestuarii (Brachyura, Carcinidae) and Pachygrapsus marmoratus (Brachyura, Grapsidae), across the Mediterranean Sea, reflect residual effects of different evolutionary histories[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-024-2401-1
Population genetic polymorphism and structure in the two coastal crab species Carcinus aestuarii (Brachyura, Carcinidae) and Pachygrapsus marmoratus (Brachyura, Grapsidae), across the Mediterranean Sea, reflect residual effects of different evolutionary histories
Laboratory of Human Genetics (LR99ES10), Faculty of Medicine of Tunis, University Tunis El Manar, Tunis 1068, Tunisia
2.
Research Unit of Genomics, Biotechnology and Antiviral Strategies (UR17ES30), University of Monastir, Higher Institute of Biotechnology of Monastir, Monastir 5000, Tunisia
3.
Laboratory of Genetics, Biodiversity and Enhancement of Bioresources (LR11ES41), University of Monastir, Higher Institute of Biotechnology of Monastir, Monastir 5000, Tunisia
4.
Instituto de Ciencias Marinas de Andalucía (ICMAN, CSIC), Campus Universitario Río San Pedro, Puerto Real, Cádiz 11519, Spain
5.
Zoology and Evolutionary Biology, University of Regensburg, Regensburg D-93040, Germany
The present investigation aims at unveiling the main causes of the recorded disparate phylogeographic patterning among the two highly dispersive coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus in the Mediterranean Sea. For this purpose, available mitochondrial and nuclear data for both species were re-analyzed and investigated for genetic polymorphism and differentiation patterns across three defined geographic scales in their distribution ranges, but also across the same locations in the Mediterranean Sea. The temporal frame of genetic diversification was also determined for both species in order to check whether observed differences in phylogeographic patterns among these coastal decapods could be attributed to different evolutionary histories. The obtained results revealed a more variable and diversified gene pool in the green crab C. aestuarii than the one recorded in the marbled crab P. marmoratus. Lack of significant correlation between pairwise genetic dissimilarities observed among C. aestuarii populations and those detected for P. marmoratus was notably discerned across the same defined Mediterranean locations. This finding indicates that the pattern of pairwise genetic differentiation does not vary in the same way in both examined crab species. Significant outputs of population genetic differentiation, retrieved within both species, were shown to be differently associated with the potential effects of various kinds of isolation processes (related to geography, environment and biogeographic boundary). Evolutionary history reconstruction showed older genetic diversification event in C. aestuarii than the one recorded in P. marmoratus. These recorded temporal frames suggest different modes of genetic diversification in both crab species (glacial vicariance for C. aestuarii and interglacial dispersal for P. marmoratus). They may also provide an explanation for the recorded differences in variation of patterns of population genetic diversity and structure, when integrated with species ecological requirements and life history traits.
The Mediterranean Sea displays one of the world's richest biological diversities. It has been considered a biodiversity hotspot region with high endemism (Coll et al., 2010). As such, it has been subjected to intense scientific research investigations. The high biodiversity and rate of endemism, recorded in the Mediterranean Sea, are postulated to be shaped by the interplay between impacts of complex geological history and particular environmental and oceanographic features (Patarnello et al., 2007; Pérez-Losada et al., 2007; Carreras et al., 2020).
The closure of the Gibraltar Strait, leading to the desiccation and isolation of the Mediterranean Sea during the Messinian Salinity Crisis (between 5.59 and 5.3 million years ago; Hsüetal., 1977), followed by the Pleistocene glacial and interglacial climate oscillations, have been considered marked instances in the biogeographic history of the Mediterranean Sea. These successive historical events have been triggering a complex chain of evolutionary events that led to eventual speciation within Atlantic-Mediterranean biota (Roman and Palumbi, 2004; Reuschel et al., 2010; González-Castellano et al., 2020). They have also left strong footprints in the contemporary genetic variability and structure of marine populations (Wilson and Eigenmann Veraguth, 2010), and influenced present distribution of Mediterranean marine species (Patarnello et al., 2007; Pérez-Losada et al., 2007). Particular regional circulation patterns within the Mediterranean Sea, acting together with the aforementioned historical isolating processes, have been also shown to play a crucial role in disrupting gene flow and shaping Mediterranean biodiversity (Duran et al., 2004; Mejri et al., 2009; Reuschel et al., 2010; Borrero-Pérez et al., 2011; Deli et al., 2016a, 2018; Pannacciulli et al., 2017). These oceanographic features include the Almeria-Oran oceanographic front, the oceanic transition between the Western and the Eastern Mediterranean (caused by a unidirectional water flow at the Siculo-Tunisian Strait and mesoscale features in the Ionian Sea (Millot and Taupier-Letage, 2005)), the contemporary hydrographic isolations of the Adriatic, Ionian and Aegean seas (maintained by a quasi-circular anticyclonic front located to the southwest of the Peloponnese Peninsula (Millot, 2005)), and the particular oceanographic features between the Marmara and Aegean seas (due to the low salinity surface water mass originating from the Black Sea).
The presence of taxonomic groups, broadly distributed throughout the Mediterranean Sea, suggests a shared biogeographic history. For instance, the interplay between impacts of palaeoenvironmental or palaeogeographic changes and contemporary hydrographical barriers are expected to shape similarly the gene pool and genetic makeup of Mediterranean marine biota. Nevertheless, recent studies found disparate patterns of population genetic structure of marine species across postulated biogeographic barriers in the Mediterranean Sea. These trends ranged from panmixia (Lourenço et al., 2017; Deli et al., 2019) to mild genetic differentiation (Fratini et al., 2016; Evangelisti et al., 2017), accentuated in certain cases to strong divergence among potential cryptic species (Reuschel et al., 2010; Weiss et al., 2018; González-Castellano et al., 2020). The recorded differences in phylogeographic patterns, among Mediterranean marine species, have often been attributed to subtle differences in ecological requirements, life-history traits (i.e., dispersal potential) and/or the evolutionary histories of individual species (Castilho et al., 2017; Catarino et al., 2017). It has been postulated that species with contrasting ecological requirements could exhibit disparate evolutionary histories and phylogeographic structures (Riginos et al., 2011). As such, a given geographic barrier may not have the same effect on all species, since life-history traits may interact differently with environmental features, determining to what extent a potential barrier will affect dispersal and gene flow. In light of these considerations, one of the main questions in evolutionary biology remains to clarify the interplay between impacts of evolutionary history and eco-biological requirements on driving disparate phylogeographic patterning within species having similar distribution ranges, and undergoing similar historical and contemporary abiotic conditions across the Mediterranean Sea.
The coastal crab species Carcinus aestuarii (Nardo, 1847) and Pachygrapsus marmoratus (Fabricius, 1787) are two of the most widely and commonly distributed species in the Mediterranean Sea. They have high fecundity and larval dispersal potential. Females of C. aestuarii produce up to 370000 embryos/brood (Grosholz and Ruiz, 2002). The fecundity estimates for P. marmoratus ranged from 8989 to 151578 eggs/crab (Aydin et al., 2014). Within C. aestuarii, the planktonic larval stage duration of approximately six weeks (Darling et al., 2008; Marino et al., 2011) is comprised of four planktonic zoeal stages (Rice and Ingle, 1975). For P. marmoratus, planktonic larval stages (corresponding to six zoeal stages) last approximately four weeks in the water column (Cuesta and Rodríguez, 2000). The spawning period of C. aestuarii occurs mainly from November to February (Mori et al., 1990). Females of P. marmoratus produce multiple broods from late spring to late summer (Flores et al., 2002). These two crab species are known to inhabit different ecological niches (with P. marmoratus prevailing in rocky shores and C. aestuarii usually found in estuarine habitats) (Mori et al., 1990; Cannicci et al., 1999; Behrens Yamada and Hauck, 2001). However, due to their wide ecological tolerance, both decapod species can be sympatric across Western and Eastern Mediterranean sites (Deli, pers. obs.).
Provided that Carcinus aestuarii and Pachygrapsus marmoratus have similar distribution ranges in the Mediterranean Sea, the postulated biogeographic boundaries in the region are expected to affect the two species in similar ways. Nevertheless, recent population genetic investigations, focusing on each species separately, unraveled different patterns of phylogeographic structure. Within C. aestuarii, mitochondrial genetic divergence between Western and Eastern Mediterranean populations was unveiled across the Siculo-Tunisian Strait (Marino et al., 2011; Ragionieri and Schubart, 2013; Deli et al., 2016a). This pattern of genetic differentiation was confirmed at the nuclear level (by means of the analysis of nuclear microsatellite loci) (Deli et al., 2016a). The occurrence of three deep mitochondrial evolutionary lineages within C. aestuarii not only accounted for a vicariant Pleistocene event across the Siculo-Tunisian Strait but also confirmed a marked phylogeographic break within the Eastern Mediterranean (Deli et al., 2016a, 2018). Against this background, lack of population genetic differentiation within P. marmoratus was unveiled among the Western and Eastern Mediterranean Basins at both mitochondrial (Cox1 gene, Fratini et al., 2011; Deli et al., 2016b) and nuclear (microsatellite loci, Deli et al., 2016b) levels. In particular, regional patterns of chaotic nuclear genetic patchiness were always evidenced within P. marmoratus across different parts of the Mediterranean Sea (Fratini et al., 2011, 2013; Deli et al., 2016b; Iannucci et al., 2020). Significant phylogeographic structure was only discerned in P. marmoratus when investigating the whole distribution range of the species, resulting in the identification of three genetically differentiated groups (East Atlantic, Mediterranean Sea, and Black Sea; Fratini et al., 2016). Noticeable concordant mitochondrial and nuclear divergence was recorded between Eastern Mediterranean and Black Sea populations of P. marmoratus across the Turkish Strait Systems (Çetin et al., 2022).
In light of these insights we asked the following main questions. First, given the different patterns of phylogeographic structure recorded among Carcinus aestuarii and Pachygrapsus marmoratus across their distribution range, do patterns of population genetic diversity and differentiation also vary differently among both species when surveyed across the same defined geographic scales? Second, provided that both species, with quite similar life history traits (i.e., high larval dispersal potential and fecundity), have experienced the same impacts of palaeogeographic and palaeoclimatic fluctuations in the Mediterranean, and are experiencing similar effects of contemporary abiotic features (i.e., environmental gradients and oceanographic patterns), why did these pressures not translate into similar patterns of population genetic structuring across main biogeographic boundaries in the Mediterranean Sea? In this context, how did both species respond to the impact of past climate fluctuations?
In order to answer these questions, both available mitochondrial and nuclear data in both species were re-analyzed and investigated for genetic polymorphisms and differentiations patterns across defined geographic scales in their distribution ranges. These data were also examined across the same locations in the Mediterranean Sea for assessment of the responses of both crab species to the impact of historical and contemporary isolating abiotic factors. Aiming to check whether observed differences in phylogeographic patterns among these coastal decapod species, with quite similar dispersal potential, could be attributed to subtle differences in the evolutionary histories of individual species, the temporal frame of genetic diversification of both species was reconstructed, based on the analysis of the mitochondrial cytochrome oxidase subunit 1 (Cox1). In this context, we hypothesize that if disparate trends of historical origin of genetic diversification turn out to be discerned, the population turnover mechanism (Grant and Bringloe, 2020) could be among many processes likely involved in driving such noticeable contrasting patterns of population genetic structure.
2.
Materials and methods
2.1
Mitochondrial and nuclear data selection
Mitochondrial (Cox1 gene) and nuclear (microsatellite loci) datasets of Carcinus aestuarii and Pachygrapsus marmoratus were retrieved from previously published population genetic investigations of both coastal crab species in their corresponding distribution areas, and were considered for the comparative analyses of genetic polymorphism and structure. Selection of the total Cox1 gene sequence dataset, corresponding to each studied crab species, was determined following coverage of much of the sequence genetic diversity (hence avoiding datasets with short sequence length), while surveying almost the entire geographic distribution of each decapod species. Taking into account these two criteria, the examined Cox1 sequences of C. aestuarii were retrieved from the previous phylogeographic investigations of Ragionieri and Schubart (2013) and Deli et al. (2016a, 2018). The already available datasets published by Darling et al. (2008) and Marino et al. (2011) were not suitable for the analysis, owing to the relatively short sequence length that may lead to the loss of considerable genetic diversity (after carrying out some preliminary analyses). The geographic spectra covered by these latter two investigations have been re-surveyed in the studies of Ragionieri and Schubart (2013) and Deli et al. (2016a, 2018). Hence, we made sure that lack of these remarkably short sequences would not affect the comparative results on population genetic diversity and differentiation. Regarding P. marmoratus, the overall dataset published by Fratini et al. (2016), including the already published sequences by Fratini et al. (2011) and Deli et al. (2016b), was implemented in the study. Recently published Cox1 sequences by Çetin et al. (2022) were not considered, provided that the resulting trimmed sequences allowed for the loss of much sequence variability (about 100 basepairs (bp) after conducting preliminary analysis). Furthermore, the geographic spectrum surveyed in the study of Çetin et al. (2022), encompassing the Eastern Mediterranean Basin and Black Sea, has been already covered and analyzed in the previous investigation of Fratini et al. (2016). In light of these considerations, a total of 412 Cox1 sequences with 519 bp length and 587 Cox1 sequences with 596 bp length were selected for C. aestuarii and P. marmoratus, respectively. In order to obtain a comprehensive and unbiased comparison, the two identified Cox1 sequences datasets for both crab species were aligned, by means of Clustal W, included in BIOEDIT (Hall, 1999), allowing for a similar fragment size analysis. As such, a final trimmed sequence length of 519 bp was finally considered for the subsequent final statistical analyses in both species. For the nuclear microsatellite loci, the generated North-African datasets for both crab species by Deli et al. (2016a, 2016b) were re-examined in this study. Despite the similarity in the number of the genotyped specimens across the African Mediterranean coast (110 individuals for each species), genotyping was carried out for five nuclear microsatellite loci within C. aestuarii and four loci within P. marmoratus. A preliminary analysis comparing two kinds of datasets (initial datasets (comprising five loci for C. aestuarii and four loci for P. marmoratus) vs. adjusted datasets (including similar number of genotyped loci for both species: four loci)) was not found to influence the outcomes of comparative genetic diversity and differentiation among both crab species (data not shown). This preliminary insight indicates that the slight difference in the number of genotyped loci within both species could not influence the results. Hence, we proceeded with the initial generated nuclear data (within both species) for the comparative estimation of population genetic polymorphism and structure variation. Details on the selected populations, number of samples per location, and study of origin, for both examined molecular (mitochondrial and nuclear) markers in both crab species, are shown in Table 1.
Table
1.
Information on the examined molecular marker, the defined geographic scale, and the selected populations (including the number of samples) for the two coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus (Further details on the geographic origin of the surveyed sampling sites for both decapods species, including countries and basins, are indicated in the references as listed within the Table)
Population
Carcinus aestuarii
Population
Pachygrapsus marmoratus
Cox1 gene
Microsatellite loci
Cox1 gene
Microsatellite loci
Micro-georaphic scale (total number = 110)
Meso-georaphic scale (total number = 133)
Macro-georaphic scale (total number = 412)
Micro-georaphic scale (total number = 90)
Meso-georaphic scale (total number = 110)
Micro-georaphic scale (total number = 60)
Meso-georaphic scale (total number = 80)
Macro-georaphic scale (total number = 587)
Micro-georaphic scale (total number = 90)
Meso-georaphic scale (total number = 110)
Tabarka
10 (a)
10 (a)
10 (a)
10 (a)
10 (a)
Tabarka
02 (d)
02 (d)
02 (d)
10 (d)
10 (d)
Bizerte
15 (a)
15 (a)
15 (a)
10 (a)
10 (a)
Bizerte
06 (d)
06 (d)
06 (d)
10 (d)
10 (d)
Sidi Rais
11 (a)
11 (a)
11 (a)
10 (a)
10 (a)
Korbos
09 (d)
09 (d)
09 (d)
10 (d)
10 (d)
Kelibia
10 (a)
10 (a)
10 (a)
10 (a)
10 (a)
Kelibia
06 (d)
06 (d)
06 (d)
10 (d)
10 (d)
Benikhiar
12 (a)
12 (a)
12 (a)
10 (a)
10 (a)
Benikhiar
06 (d)
06 (d)
06 (d)
10 (d)
10 (d)
Monastir
11 (a)
11 (a)
11 (a)
10 (a)
10 (a)
Monastir
06 (d)
06 (d)
06 (d)
10 (d)
10 (d)
Chebba
12 (a)
12 (a)
12 (a)
10 (a)
10 (a)
Chebba
05 (d)
05 (d)
05 (d)
10 (d)
10 (d)
Sfax
15 (a)
15 (a)
15 (a)
10 (a)
10 (a)
Sfax
09 (d)
09 (d)
09 (d)
10 (d)
10 (d)
Djerba
14 (a)
14 (a)
14 (a)
10 (a)
10 (a)
Zarzis
11 (d)
11 (d)
11 (d)
10 (d)
10 (d)
Tripoli
11 (a)
11 (a)
10 (a)
Tajura
10 (d)
10 (d)
10 (d)
Mosrata
12 (a)
12 (a)
Annaba
10 (d)
10 (d)
Ebro Delta
28 (b)
Terciera
17 (f)
Camargue
20 (b)
Fuerteventura
18 (e)
Livorno
16 (b)
Sesimbra
18 (e)
Elba
16 (b)
Cala Iris
15 (e)
Sardinia
18 (b)
Valencia
20 (e)
Lago Lungo
15 (b)
Tarragona
20 (e)
Fusaro
15 (b)
Saint Florent
15 (e)
Sicily
10 (b)
Calafuria
15 (e)
Venice Lagoon
15 (a, b)
Porto di Follonica
05 (f)
Pomer
20 (b)
Porto Ercole
15 (e)
Amvrakikos
14 (b, c)
Giglio
11 (e)
Lefkada
14 (c)
Rocchette
14 (e)
Navarino
10 (c)
Montecristo
15 (e)
Peloponnesus
06 (b)
Gorgona
09 (f)
Alexandroupolis
12 (c)
Gaeta
15 (e)
Izmir Bay
06 (c)
Fusaro
15 (e)
Enez Dalyan Lagoon
20 (c)
Crotone
16 (e)
Dardanelles Strait
05 (c)
Messina
16 (e)
Prince’s Islands
12 (c)
Porto Cesareo
07 (f)
Bosphorus Strait
07 (c)
Othonoi
07 (f)
Ancona
07 (f)
Otranto
07 (f)
Torre Guaceto
07 (f)
Tremiti
07 (f)
Karaburun Peninsula
07 (f)
Kornati
07 (f)
Pula
18 (f)
Zadar
11 (f)
Boka Kotorska Bay
07 (f)
Crete
14 (f)
Paros Naoussa harbour
07 (f)
Chalkidiki
08 (f)
Beldibi
18 (e)
Girne
22 (f)
Sile
07 (f)
Sinop
07 (f)
Sozopol
21 (f)
Cape Kaliakra
07 (f)
Ropotamo-Kiten
07 (f)
Batumi
07 (f)
Karadag
07 (f)
Tarhankut
07 (f)
Costinesti
07 (f)
Note: (a) Deli et al. (2016a); (b) Ragionieri and Schubart (2013); (c) Deli et al. (2018); (d) Deli et al. (2016b); (e) Fratini et al. (2011); (f) Fratini et al. (2016). The micro-geographic scale corresponds to the Tunisian coast. The meso-geographic scale refers to the North-African coast. It is defined by either the Tunisian and Libyan littorals for the Cox1 data of Carcinus aestuarii or the Tunisian, Algerian and Libyan coasts for the Cox1 data of Pachygrapsus marmoratus. For the examined microsatellite data of both crab species, the meso-geographic scale is represented by the Tunisian and Libyan littorals. The macro-geographic sale corresponds to the distribution range of each examined crab species.
2.2.1
Assessments of geographic variation of population genetic polymorphism and differentiation
Patterns of genetic polymorphism variation in the two studied crab species were assessed and compared across three defined geographic scales (Table 1). The micro-geographic scale corresponds to the Tunisian coast. The meso-geographic scale refers to the North-African coast. It is defined by either the Tunisian and Libyan littorals for the Cox1 data of Carcinus aestuarii or the Algerian, Tunisian and Libyan coasts for the Cox1 data of Pachygrapsus marmoratus. For the examined microsatellite data of both crab species, the meso-geographic scale encompasses the Tunisian and Libyan coast. Although the micro- and meso-geographic scales do not differ greatly in their geographic extension, the choice of these scales is justified by the fact that the micro-geographic scale encompasses the biogeographic transition zone at the Siculo-Tunisian Strait while the meso-geographic scale includes both the Siculo-Tunisian Strait and the Tunisian-Libyan Gulf region (known to harbor a historical glacial refugium (Deli et al., 2016a)). The macro-geographic scale corresponds to the entire distribution range of each examined crab species. Three parameters of mitochondrial diversity (haplotype diversity, nucleotide diversity, and mean number of nucleotide differences) and three indices of nuclear diversity (expected heterozygosity, allelic richness, and mean number of alleles per locus) were used for the comparative analyses. Mitochondrial diversity variation has been assessed and compared among both species across the three defined geographic scales. Measures and comparisons of nuclear genetic diversity, by means of the nuclear microsatellite loci, were carried out only across the micro- and meso-geographic scales. In order to obtain a comprehensive and an unbiased inference of comparison of genetic polymorphism among the two studied species, across the three defined geographic scales, we used the same number of specimens, sequenced for the Cox1 gene and genotyped for nuclear microsatellite loci, for each kind of assessment and comparison across a given geographic spectrum. Provided that earlier studies by Deli et al. (2016a, 2016b) obtained more Cox1 sequences for C. aestuarii than P. marmoratus across the micro-geographic scale of the Tunisian coast, but also across the meso-geographic scale encompassing the North-African coast, we applied a rarefaction procedure in order to discard the possible bias of sample size on inferred genetic diversity. For this purpose, we used the number analyzed for P. marmoratus (corresponding to the size of the smallest dataset) as threshold for comparison (60 examined sequences of Cox1 gene from the Tunisian coast and 80 generated Cox1 sequences from the North-African coast). For each case of comparison, a subsample of 60 and 80 individuals (characterizing the Tunisian micro-geographic scale and the North-African meso-geographic spectrum respectively) from both species were drawn randomly, and genetic diversity indices were calculated. In order to infer reliable statistics, the procedure was repeated at least 50 times for each sample, and then the mean subsample diversity for each sample was calculated. This procedure has been similarly adopted for genetic polymorphism comparison among both crab species across the macro-geographic scale. For this kind of analysis, a subsample of 412 individuals (corresponding to the size of the smallest dataset of C. aestuarii) was implemented in the rarefaction procedure as already described. Noteworthy, this strategy was not applied for microsatellite loci provided the equal number of specimens genotyped for these markers for each species across each surveyed geographic scale (90 specimens from the Tunisian coast and 110 samples from the North-African coast; Table 1).
Using the same dataset, surveyed already for inter-species genetic polymorphism comparison, overall genetic differentiation, reflecting population genetic structure, was assessed for each examined molecular marker, across each defined geographic scale, by means of one-level analysis of molecular variance (AMOVA) (Excoffier et al., 1992), as implemented in ARLEQUIN version 3.1 (Excoffier et al., 2007). This genetic distance measurement was inferred from both nucleotide divergence (based on the Tamura-Nei model (Tamura and Nei, 1993), equivalent to the nucleotide substitution models determined for both crab species by the software MEGA X (Kumar et al., 2018) using the Find best DNA models [ML] option in MEGA X (Nei and Kumar, 2000; Tamura et al., 2011), see below for further details) and haplotype frequencies for the mtDNA Cox1 gene. For microsatellite loci, the distance method, based on the number of different alleles, included in ARLEQUIN version 3.1, was selected to carry out the analyses across the micro- and meso-geographic scales. For both analyzed genetic markers, significant trends of overall genetic differentiation (P<0.05), across the various geographic scales, were assessed by a randomization procedure with 10000 permutations. As has been considered for comparative genetic polymorphism analyses, the same rarefaction procedure was applied to infer estimates of overall genetic differentiation within both studied crab species across the different surveyed geographic scales.
2.2.2
Analysis of correlation between trends of population genetic structure across the Mediterranean Sea, and assessment of the responses of both crab species to the impact of historical and contemporary isolating abiotic factors
In order to assess correlation between trends of population genetic structure discerned in Carcinus aestuarii and Pachygrapsus marmoratus, and see whether patterns of genetic differentiation could vary similarly in the two crab species, analyses were conducted across the same sampling sites of occurrence (or across those very geographically proximate) within the Mediterranean Sea (based on the studies of Fratini et al. (2011, 2016), Ragionieri and Schubart (2013), and Deli et al. (2016a, 2016b, 2018)). This latter geographic area within the distribution area of both species was selected based on the facts that C. aestuarii has not an East Atlantic distribution, and samples of this species are still lacking from the Black Sea (Deli et al., 2018). For this purpose, a set of 17 sampling locations, assigned to the Western and Eastern Mediterranean Basins, were chosen (Fig. 1, Table 2). Overall, the selection of the 17 sites targeted for mitochondrial Cox1 data analyses, corresponding to the same geographic origin or representing geographically very close locations, resulted in defining a total of 245 Cox1 sequences for C. aestuarii and 189 Cox1 sequences for P. marmoratus. For the nuclear microsatellite data, the already examined ten African Mediterranean locations for both crab species (Deli et al., 2016a, 2016b), spanning the Western and Eastern Mediterranean Basins, were used for the analysis (a total of selected genotyped 110 specimens for each examined species) (Fig. 1, Table 2). The relationship between average genetic dissimilarities (based on F-Statistics: ΦST and FST for the mitochondrial Cox1 gene and FST for the nuclear microsatellite loci) among populations of C. aestuarii and those detected among P. marmoratus populations was assessed using the Linear, one independent, n dependent (multivariate regression) model, implemented in the software PAST version 2.17 (Hammer et al., 2001). This model can fit each dependent variable separately to the independent variable using simple linear regression. For each examined molecular marker, the dataset obtained for C. aestuarii was considered as an independent variable and the corresponding one for P. marmoratus as a dependent variable. The “adjusted” correlation coefficient R2 associated with an overall MANOVA test of multivariate regression significance were provided.
Figure
1.
Geographic distribution of the selected seventeen sampling locations harbouring the coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus across the Mediterranean Sea. Details on these localities, including the corresponding number of analyzed samples of each species per site, were highlighted in Table 2. S-T S represents the Siculo-Tunisian Strait. Construction of the base map was carried out by the software DIVA-GIS 7.5.0 (http://www.diva-gis.org).
Table
2.
Information on the selected (same or geographically very close) Mediterranean sites harboring the two coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus (examined for mitochondrial and nuclear markers), regions, and the number of samples per location
Selected (same or geographically very close) Mediterranean sites harbouring both species
Region
Cox1 gene
Microsatellite loci
Carcinus aestuarii
Pachygrapsus marmoratus
Carcinus aestuarii
Pachygrapsus marmoratus
Location
Sample number
Location
Sample number
Location
Sample number
Location
Sample number
Site (1)
Western Mediterranean
Ebro
28
Tarragona
20
Site (2)
Western Mediterranean
Livorno
16
Calafuria
15
Site (3)
Western Mediterranean
Elba
16
Porto di Follonica + Rocchette*
19
Site (4)
Western Mediterranean
Lago Lungo
15
Gaeta
15
Site (5)
Western Mediterranean
Fusaro
15
Fusaro
15
Site (6)
Western Mediterranean
Tabarka
10
Tabarka + Annaba*
12
Tabarka
10
Tabarka
10
Site (7)
Western Mediterranean
Bizerte
15
Bizerte
06
Bizerte
10
Bizerte
10
Site (8)
Western Mediterranean
Sidi Rais
11
Korbos
09
Sidi Rais
10
Korbos
10
Site (9)
Western Mediterranean
Kelibia
10
Kelibia
06
Kelibia
10
Kelibia
10
Site (10)
Western Mediterranean
Benikhiar
12
Benikhiar
06
Benikhiar
10
Benikhiar
10
Site (11)
Eastern Mediterranean
Monastir
11
Monastir
06
Monastir
10
Monastir
10
Site (12)
Eastern Mediterranean
Chebba
12
Chebba
05
Chebba
10
Chebba
10
Site (13)
Eastern Mediterranean
Sfax
15
Sfax
09
Sfax
10
Sfax
10
Site (14)
Eastern Mediterranean
Djerba
14
Zarzis
11
Djerba
10
Zarzis
10
Site (15)
Eastern Mediterranean
Tripoli
11
Tajura
10
Tripoli
10
Tajura
10
Site (16)
Eastern Mediterranean
Pomer
20
Pula
18
Site (17)
Eastern Mediterranean
Amvrakikos
14
Othonoi
07
Note: * These geographically very close sites were combined for Pachygrapsus marmoratus in order to increase the sample size.
In order to determine whether both species responded similarly to the impact of historical and contemporary isolating abiotic factors, significant outputs of overall population genetic differentiation (corresponding to the selected Mediterranean dataset), inferred from one-level AMOVA, were assessed for the effect of isolation by distance (IBD), isolation by environment (IBE), and isolation by resistance (IBR) across the selected geographic area within the Mediterranean Sea. Unlike IBD, both IBE and IBR were newly investigated for both species in this study. For the IBD test, correlation between pairwise genetic and geographic distances was assessed by means of the Mantel test (Mantel, 1967), as implemented in ARLEQUIN version 3.1. A total of 10000 random permutations were run in order to assess the statistical significance of the test. In order to discern evidence for IBE, we extracted data on sea surface temperature and salinity for each examined location. Sea surface temperatures of the explored Mediterranean sites were obtained from the two databases at http://www.seatemperature.org and http://www.worldseatemp.com. Sea surface salinities were estimated from the Integrated Climate Data Center at http://icdc.cen.uni-hamburg.de/1/daten/ocean/smos-sss.html. The obtained mean values of sea surface temperature, averaged across the twelve months, and those of salinity were then subjected to principal component analysis (PCA) using the software PAST version 2.17 (Hammer et al., 2001). The first two principal components were used as points to calculate a pairwise environmental distance matrix for all populations, and IBE (correlation between pairwise genetic and environmental distances) was assessed for each kind of examined (mitochondrial and nuclear) dataset within both studied crab species using ARLEQUIN. Aiming at determining whether the obtained population genetic differentiation could be linked to the impact of the barrier to gene flow at the Siculo-Tunisian Strait, we subsequently tested the IBR hypothesis. For such kind of assessment, the selected Mediterranean populations were divided into two main groups in each species, east and west of the Siculo-Tunisian Strait (Eastern Mediterranean and Western Mediterranean populations as already defined). Pairs of populations separated by this biogeographic boundary (those corresponding to the Eastern and Western Mediterranean) were coded 1, while pairs of populations assigned to the same Mediterranean Basin (corresponding either to the Eastern Mediterranean or Western Mediterranean) were coded 0. Correlation between this designed distance matrix and genetic distances was assessed in ARLEQUIN. Similarly to the IBD test, statistical significances of the IBE and IBR tests were assessed by running a total of 10000 random permutations. It needs to be noted that, for the mitochondrial Cox1 marker, pairwise genetic distances, used for the three aforementioned correlations, were computed from the Tamura and Nei distance as the overall genetic differentiation (based on this latter distance) was shown to exceed the one estimated from haplotype frequencies.
2.2.3
Estimation of genetic diversification times within both crab species
In order to assess the impact of evolutionary history on patterns of genetic polymorphism and structuring variation in Carcinus aestuarii and Pachygrapsus marmoratus, temporal patterns of genetic diversification were determined in both species. The same length of Cox1 dataset alignment, as previously determined (519 bp) for the two coastal crab species, was implemented in the analysis in order to infer comprehensive comparison. Estimation of genetic diversification times within C. aestuarii and P. marmoratus was determined with the software package BEAST version 1.7.5 (Drummond et al., 2012). Prior to BEAST analyses, the optimal nucleotide substitution model, best fitting each examined dataset, was selected by means of the software MEGA X (Kumar et al., 2018) using the Find best DNA models (ML) option in MEGA X (Nei and Kumar, 2000; Tamura et al., 2011). Notably, appropriate calibrations or substitution rates were applied in order to avoid overestimation of diversification times when implementing unspecific published mutation rates related to other decapods genera or families. For C. aestuarii, the specifically estimated Cox1 gene mutation rate for Carcinus (3.86% per million years, Myr; Marino et al., 2011) was used to calibrate the genealogy and date the time to the most recent common ancestor (TMRCA) of the examined Cox1 dataset. Regarding P. marmoratus, we determined a mutation rate for the Cox1 mitochondrial gene in a species-level phylogenetic analysis, involving the highly diversified Cox1 dataset (a total of 154 haplotypes) recovered in the relatively closely related species Pachygrapsus crassipes across the northeastern and western Pacific coasts (Cassone and Boulding, 2006; GenBank accession numbers: AY951986-AY952011 and AY952012-AY952110) along with P. marmoratus haplotypes inferred from the defined dataset in this study. We used the estimated time of divergence (between 0.8 and 1.2 million years ago) among the two trans-Pacific lineages identified in P. crassipes (corresponding to the Eastern and Western Pacific) (Cassone and Boulding, 2006) as a calibration point for mutation rate estimation within Pachygrapsus sp.. The software BEAST version 1.7.5 (Drummond et al., 2012) was used to determine the Cox1 substitution rate. The retrieved TN93+G+I model was found to fit the analyzed dataset well (exhibiting the lowest Bayesian Information Criterion (BIC) score, according to the outcome of Maximum Likelihood fits of 24 different nucleotide substitution models tested by MEGA X). We used the two parameter Birth-Death model, regarded as an appropriate tree prior for species diversification (Nee et al., 1994), and implemented an uncorrelated log-normal relaxed clock. A total run of 30 million generations were specified for the Markov chain Monte Carlo (MCMC) simulations. The software TRACER version 1.5 (Rambaut and Drummond, 2009) was used to check the convergence of the runs (effective sample sizes, ESS, of all parameters greater than 200) and exhibit the result corresponding to the mean mutation rate (and its 95% high posterior density interval (HPD)).
Subsequently, a set of tree and molecular clock models were tested for the reliable and comprehensive inference of the temporal frame of genetic diversification within the studied crab species Carcinus aestuarii and Pachygrapsus marmoratus. For both taxa, the coalescent Bayesian Skyline and the strict molecular clock models were selected as priors for the BEAST analyses. The HKY+G+I model, identified as the most appropriate model of sequence evolution by MEGA for C. aestuarii Cox1 haplotype dataset, was implemented in the BEAST analysis. For P. marmoratus, the TN93+G model, determined as the best fitting nucleotide substitution model, was incorporated in the BEAST analysis. Both already estimated and newly determined Cox1 mutation rates for C. aestuarii and P. marmoratus, respectively, were used to calibrate the haplotype phylogeny and estimate the TMRCA in the Cox1 dataset of each species. MCMC simulations and sampled steps were appropriately set according to each examined Cox1 dataset (100 million generations sampled every 1000 generations for C. aestuarii, and 60 million generations sampled every 1000 generations for P. marmoratus) in order to reach convergence of the runs (effective sample sizes, ESS, of all parameters greater than 200) checked by TRACER version 1.5. For each analyzed dataset, the first 20% of sampled genealogies were discarded as burn-in, and the maximum clade credibility tree was computed with TreeAnnotator version 1.7.5 (Drummond et al., 2012). The obtained calibrated Cox1 haplotype genealogy, for each species, was exhibited and checked for the TMRCA in FigTree version 1.4.0 (Rambaut, 2012).
3.
Results
3.1
Inter-species comparison of genetic polymorphism and overall genetic differentiation variations across the surveyed geographic scales
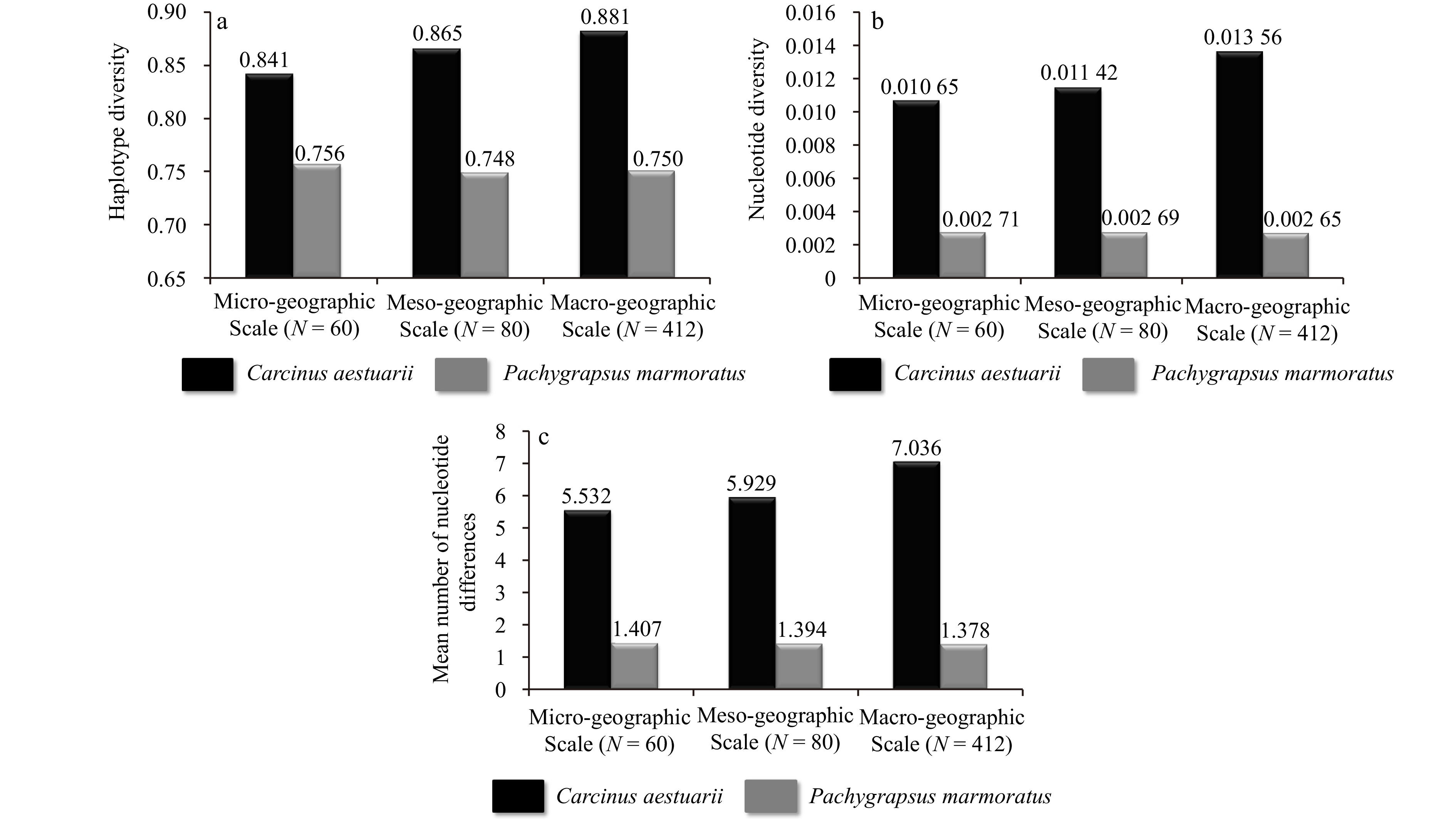
Comparison of genetic diversity parameters, corresponding to the mitochondrial Cox1 gene and nuclear microsatellite loci, between Carcinus aestuarii and Pachygrapsus marmoratus, showed marked difference in terms of genetic polymorphism level and variation across the three surveyed geographic scales in this study. When analyzing and comparing the same Cox1 length for genetic polymorphism variation among the same number of sequences in both species, remarkably higher levels of mitochondrial genetic diversity, in terms of haplotype diversity, nucleotide diversity and mean number of nucleotide differences, were unveiled within C. aestuarii than those detected in P. marmoratus (Fig. 2). Notably, the three mitochondrial parameters were found to increase proportionally with the investigated geographic area for C. aestuarii (Fig. 2). This trend of variation was not discerned in P. marmoratus (Fig. 2). Regarding the microsatellite loci, the analysis of three nuclear genetic diversity parameters (expected heterozygosity, allelic richness and mean number of alleles per locus) clearly revealed higher genetic diversity in C. aestuarii than the one recorded in P. marmoratus across the two geographic spectra corresponding to the micro-geographic scale of the Tunisian coast and the relatively enlarged meso-geographic scale encompassing the North-African (Tunisian and Libyan) Mediterranean coast (Fig. 3). Opposed to the outcome of Cox1 mitochondrial data analysis, similar outputs of variation patterns at nuclear microsatellite loci were retrieved for C. aestuarii and P. marmoratus. In particular, a very slight increase in the three examined nuclear genetic diversity indices was noticed for the two species across the surveyed geographic scales (Fig. 3).
Figure
2.
Variation trends of mitochondrial genetic polymorphism (a. haplotype diversity, b. nucleotide diversity, and c. mean number of nucleotide variation) in the two coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus across the three surveyed geographic scales (as defined in Table 1). For each geographic spectrum, levels of genetic diversity parameters were computed and inferred for the same number of specimens (N) corresponding to both examined species.
Figure
3.
Variation trends of nuclear genetic polymorphism (a. expected heterozygosity, b. allelic richness, and c. mean number of alleles per locus) in the two coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus across the two surveyed micro- and meso-geographic scales (as defined in Table 1). For each geographic spectrum, levels of genetic diversity parameters were computed and inferred for the same number of specimens (N) corresponding to both examined species.
Assessment of overall mitochondrial and nuclear population genetic differentiation unveiled significant population genetic structure in Carcinus aestuarii along the three surveyed geographic spectra (Table 3).This recorded pattern clearly contrasted with that inferred for Pachygrapsus marmoratus. Notably, for the latter species, lack of significant overall genetic subdivision, at the mitochondrial Cox1 gene, was discerned across the micro- and meso-geographic scales, suggesting the existence of panmictic gene pools at the African Mediterranean coast. When focusing on the entire distribution area of P. marmoratus, trend of population genetic structure, based on both Tamura-Nei distances and haplotype frequencies, turned out to be highly significant (Table 3). Notably, both crab species exhibited significant mitochondrial genetic structure across the defined macro-geographic scale representing the entire distribution of both taxa (Table 3). Overall genetic differentiation, at the nuclear microsatellite markers, was found to be highly significant for the two coastal crab species across the defined micro- and meso-geographic scales (Table 3). Similar to the outcomes of geographic trends of variation of genetic polymorphism, overall mitochondrial genetic differentiation within C. aestuarii (based on Tamura-Nei distances) was found to increase when enlarging the analyzed geographic spectrum (Table 3). Such noticeable variation tendency was not maintained (according to both nucleotide divergence and haplotype frequency) in P. marmoratus across the same surveyed geographic scales (Table 3). Based on the analysis of nuclear microsatellite loci, both crab species exhibited similar variation tendency of genetic differentiation. Notably, the recorded FST values were shown to slightly increase with the geographic spectrum extension (stretching to the Libyan coast) (Table 3).
Table
3.
Comparison of overall genetic differentiation among the two sympatric species Carcinus aestuarii and Pachygrapsus marmoratus across the three defined geographic scales (micro-geographic, meso-geographic, and macro-geographic scales, as indicated in Table 1)
Species/molecular marker examined for genetic differentiation across each geographic scale
Cox1 mitochondrial gene
Microsatellite nuclear loci
Micro-geographic scale (N=60)
Meso-geographic scale (N=80)
Macro-geographic scale (N=412)
Micro-g eographic scale (N=90)
Meso- geographic scale (N=110)
Distance method: Tamura and Nei distance (P)
Distance method: Haplotype frequency (P)
Distance method: Tamura and Nei distance (P)
Distance method: Haplotype frequency (P)
Distance method: Tamura and Nei distance (P)
Distance method: Haplotype frequency (P)
Distance method: number of different alleles (P)
Distance method: number of different alleles (P)
Carcinus aestuarii
ΦST=0.177 (P<0.05)
FST=0.134 (P<0.01)
ΦST=0.233 (P<0.01)
FST=0.116 (P<0.01)
ΦST=0.569 (P<0.001)
FST=0.108 (P<0.001)
FST=0.013 (P<0.01)
FST=0.016 (P<0.001)
Pachygrapsus marmoratus
ΦST=0.043 (P=0.073)
FST=0.028 (P=0.151)
ΦST=0.027 (P=0.130)
FST=0.013 (P=0.263)
ΦST=0.210 (P<0.001)
FST=0.150 (P<0.001)
FST=0.020 (P<0.001)
FST=0.022 (P<0.001)
Note: The computed values for mitochondrial genetic distances were obtained after a rarefaction procedure as described in details in the material and methods section. The values, highlighted for each genetic distance measure, were inferred from the same compared number (N) of samples representing the set of populations included in each defined geographic spectrum (as cited in Table 1). Trend and significance (P) of overall genetic differentiation were calculated from 10000 permutations. Significant values are denoted in bold.
Taking into account all of these aforementioned new insights into patterns of variation of mitochondrial and nuclear genetic diversity and structure, the green crab Carcinus aestuarii seems to be genetically more variable and diversified than the marbled crab Pachygrapsus marmoratus.
3.2
Correlation between trends of population genetic differentiation within both species across the Mediterranean Sea, and assessment of potential impacts of geography, environment and historical biogeographic boundary
The multivariate regression analysis revealed lack of significant correlation between pairwise genetic dissimilarities observed among Carcinus aestuarii populations and those detected for Pachygrapsus marmoratus across the same defined Mediterranean locations. This finding was unveiled for the mitochondrial Cox1 gene, based on Tamura and Nei distances (R2=0.00030, P (regression)=0.843) and haplotype frequencies (R2=0.02559, P (regression)=0.063) (Fig. 4). It was also discerned for the nuclear microsatellite loci, based on the number of different alleles (R2=0.0509, P (regression)=0.136) (Fig. 4). Overall, the obtained results support the tendency that the pattern of pairwise genetic differentiation does not vary in the same way in both examined crab species across the surveyed geographic spectrum of the Mediterranean Sea.
Figure
4.
Multivariate regression analysis depicting correlation between average genetic dissimilarities (genetic distances) detected among Carcinus aestuarii populations and those discerned among Pachygrapsus marmoratus populations from the same Mediterranean locations (as indicated in Table 2 and exhibited in Fig. 1), for both mitochondrial Cox1 gene (based on Tamura and Nei distances and haplotype frequencies) and nuclear microsatellite loci (based on the number of different alleles).
Assessment of overall mitochondrial genetic differentiation within the analyzed Mediterranean dataset, based on Tamura-Nei distance, unveiled significant genetic subdivision within Carcinus aestuarii (ΦST=0.221; P<0.001) versus marked genetic panmixia within Pachygrapsus marmoratus (ΦST=0.011; P=0.223) across the same surveyed geographic area. The recorded significant population genetic differentiation within C. aestuarii was shown to be potentially driven by isolation by environment and isolation by resistance, but not by isolation by distance (Table 4).
Table
4.
Investigation of the origin of genetic differentiation within sympatric Mediterranean populations of Carcinus aestuarii and Pachygrapsus marmoratus (as indicated in Table 2 and shown in Fig. 1), highlighting patterns of overall genetic differentiation based on the analyses of mitochondrial and nuclear genetic markers and assessing correlations (R) between genetic distances and geographic distances (isolation by distance), environmental distances (isolation by environment), and the distance matrix defined according to the potential influence of the biogeographic boundary at the Siculo-Tunisian Strait (isolation by resistance)
Studied species/Analyzed molecular marker for potential origins of genetic differentiation
Carcinus aestuarii
Pachygrapsus marmoratus
Mitochondrial DNA (Cox1 gene)
Nuclear DNA (microsatellite loci)
Mitochondrial DNA (Cox1 gene)
Nuclear DNA (microsatellite loci)
Implemented genetic distance (overall genetic differentiation and its significance)
Tamura-Nei distance (ΦST=0.221; P=0.000)
number of different alleles (FST=0.016; P=0.000)
Tamura-Nei distance (ΦST=0.011; P=0.223)
number of different alleles (FST=0.024; P=0.000)
Isolation by distance (IBD)
R=0.080 (P=0.352)
R=0.583 (P=0.000)
not applicable
R=0.074 (P=0.628)
Isolation by environment (IBE)
R=0.337 (P=0.000)
R=0.569 (P=0.000)
not applicable
R=−0.157 (P=0.303)
Isolation by resistance (IBR)
R=0.263 (P=0.002)
R=0.219 (P=0.146)
not applicable
R=0.010 (P=0.947)
Note: Significant correlations (P<0.05) are marked in bold. In the case of non-significant overall genetic differentiation, all of the three aforementioned kinds of correlations cannot be computed (not applicable).
At the nuclear microsatellite loci, both geographic and environmental distances were significant predictors of genetic distances for the African Mediterranean dataset of Carcinus aestuarii (Table 4). In contrast, none of the three analyzed isolating effects were involved in generating the significant pattern of genetic differentiation within Pachygrapsus marmoratus (Table 4) across the same surveyed geographic spectrum. Although being not significant, the effect of the biogeographic boundary at the Siculo-Tunisian Strait was found to be markedly higher in C. aestuarii (R=0.219) than in P. marmoratus (R=0.010).
3.3
Temporal patterns of genetic diversification within both crab species
The construction of calibrated Bayesian phylogeny of the Cox1 haplotypes dataset for each studied crab species, based on the analysis of the same aligned fragment length of the mitochondrial Cox1 gene (519 bp), yielded different temporal frames of genetic diversification. When implementing the already published Cox1 mutation rate within Carcinus (3.86% per Myr; Marino et al., 2011), diversification of the recorded haplotypes within the green crab Carcinus aestuarii was found to start approximately at 0.562 million years ago (Mya) (95% HPD: 0.340-0.801 Mya). For the marbled crab Pachygrapsus marmoratus, genetic diversification time of the examined haplotype dataset dated back to 0.129 Mya (95% HPD: 0.072-0.200 Mya) when applying the newly determined Cox1 mutation rate for Pachygrapsus (3.05% per Myr (95% HPD: 1.03%-5.84%)). These results indicate an older genetic diversification event in C. aestuarii than the one recorded in P. marmoratus.
4.
Discussion
The present investigation is the first to assess and compare patterns of population genetic polymorphism and structure variation of two coastal crab species across defined geographic scales. It allowed unraveling potential historical and contemporary abiotic drivers of population genetic subdivision for each species across the same sampling locations within the Mediterranean Sea. It also provided preliminary insights into the evolutionary history that was potentially responsible for shaping the population genetic diversity and phylogeographic structure as currently observed in both species. A more variable and diversified gene pool in the green crab Carcinus aestuarii than the one recorded in the marbled crab Pachygrapsus marmoratus was unveiled in this study. This finding was inferred from the comparison of genetic diversity parameters and differentiation metrics, corresponding to the mitochondrial Cox1 gene and nuclear microsatellite loci, between the two coastal crab species across the three surveyed geographic spectra. The obtained results also revealed lack of significant correlation between pairwise genetic dissimilarities observed among C. aestuarii populations and those detected for P. marmoratus across the same defined Mediterranean locations. This finding supports the tendency that the pattern of pairwise genetic differentiation does not vary in the same way in both examined crab species across the surveyed geographic spectrum of the Mediterranean Sea. Notably, significant outputs of population genetic differentiation, discerned within both species, were shown to be differently associated with the potential effects of the various kinds of isolation processes (IBD, IBE, and IBR). Different temporal frames of genetic diversification (based on the analysis of the same aligned fragment length of the mitochondrial Cox1 gene) indicated older genetic diversification event in C. aestuarii than the one recorded in P. marmoratus.
Inter-species assessment and comparison of genetic polymorphism showed higher levels of mitochondrial and nuclear genetic diversity in the green crab Carcinus aestuarii than those detected in the marbled crab Pachygrapsus marmoratus across the three surveyed geographic scales. This finding stemmed from the analysis of the same number of specimens, same Cox1 fragment length, and relatively same set of genotyped nuclear microsatellite loci. Furthermore, geographical trends of variation of these parameters were shown to differ among both species, specifically, at the mitochondrial Cox1 gene. Such disparate patterns of genetic polymorphism levels could be linked to the impact microevolutionary forces acting within the habitats of both species including genetic drift, selection, and gene flow. It is known that C. aestuarii inhabits estuaries and lagoons (Mori et al.,1990; Behrens Yamada and Hauck, 2001), while P. marmoratus is prevalent in rocky intertidal environment (Cannicci et al., 1999). Accordingly, the interplay between influences of the aforementioned microevolutionary forces and species life history traits (i.e., larval retention and behavior as well as ecological requirements) could act differently on gene pools in the two kinds of habitats, leading to different levels of genetic polymorphism within both examined crab species. Different evolutionary forces, such as genetic drift and natural selection, have been suggested to influence significantly patterns of genetic polymorphism in estuaries and lagoons habitats, compared to their mild effects in the seemingly homogeneous coastal rocky environments (Lévêque, 1997; Bilton et al., 2002). This insight derived from the fact that estuaries and lagoons are characterized by frequent alteration of ecological conditions over space and time (Lévêque, 1997; Bilton et al., 2002). Moreover, numerous investigations have shown that genetic connectivity among populations of estuarine taxa is more restricted than that occurring in species inhabiting the open coast (Ward et al., 1994; Watts and Johnson, 2004; Pelc et al., 2009). In light of these considerations, genetic drift is likely to influence gene pools in these relatively isolated estuarine environments by increasing inter-population genetic polymorphism. This latter assumption could explain the marked increase in levels of mitochondrial and nuclear genetic diversity within C. aestuarii along the three surveyed geographic spectra, opposed to what has been recorded in P. marmoratus. Alternatively, the different recorded contemporary patterns of genetic polymorphisms among C. aestuarii and P. marmoratus could be reflective of different evolutionary histories hinting at potential disparate responses of both species to palaeoclimate and palaeogeographic shifts during the Quaternary climate oscillations. Recent population genetic investigations on the green crab allowed identifying three historically divergent haplogroups likely assigned to three glacial refugia (Deli et al., 2016a, 2018). Among these, two haplogroups (or genetic types 2 and 3) were mainly delineated in the Tunisian-Libyan Gulf and the Aegean Sea respectively, and one haplogroup (or genetic type 1) was hypothesized to have originated in the Western Mediterranean (Ragionieri and Schubart, 2013; Deli et al., 2016a, 2018). For the marbled crab, however, shallow pattern of mitochondrial genealogy, with no pattern of deep genetic separation or geographic delineation of the recorded sub-clades (except for a relatively distinct haplogroup confined to Eastern Mediterranean and Black Sea), was postulated to reflect residual effect of recent evolutionary history of the species. This scenario involves potential dispersal events from the highly genetically diversified region of the Mediterranean Sea into the East Atlantic and Black Sea (Deli et al., 2016b; Fratini et al., 2016). Such geographic delineation of potential historical repositories of genetic diversity within the two species matches patterns of variation of mitochondrial genetic diversity across the three surveyed geographic scales. As such, it could explain the recorded disparate patterns of genetic polymorphism levels among both decapods species. It also suggests that complex evolutionary history, such as the one discerned in the green crab C. aestuarii, may be potential predictor of higher genetic diversity levels at both mitochondrial and nuclear examined markers.
Assessment of overall population genetic differentiation unraveled significant population genetic structure within Carcinus aestuarii, at the mitochondrial and nuclear levels, across the three defined geographic scales. This trend clearly contrasts with the one recorded within Pachygrapsus marmoratus. For the latter species, lack of mitochondrial population genetic structure was discerned across the micro- and meso-geographic scales. Notably, for the mtDNA Cox1 gene, estimated overall genetic distances (based on Tamura-Nei distance and haplotype frequency) were remarkably higher in the green crab species than those recorded in the marbled crab species. In contrast, nuclear genetic differentiation metrics were comparable between the two species, across the only two examined geographic spectra corresponding to the Tunisian and North-African Mediterranean coasts. The different trends of mitochondrial population genetic structure among both species, specifically at the micro- and meso-geographic scales, likely reflect the residual effects of different evolutionary histories of both species across the Siculo-Tunisian Strait with the potential joint impact of the habitat type. The study of Deli et al. (2016a) showed that specimens of the estuarine marine species C. aestuarii withstood the Pleistocene glacial climate deterioration across the Siculo-Tunisian Strait by surviving in distinct Western and Eastern Mediterranean glacial refugia. This historical isolation allowed populations from both Mediterranean regions to accumulate significant genetic differentiation in allopatry followed by secondary contact and admixture. For the intertidal rocky marine species P. marmoratus, the considerable genetic panmixia, unveiled across the same geographic area, encompassing the North African Mediterranean coast (Deli et al., 2016b), suggests that the marbled crab species responded differently to the Pleistocene climate shifts across the Siculo-Tunisian Strait. Notably, the intertidal rocky marine biota is known to be the most affected and vulnerable to the Pleistocene climate deteriorations due to the reduction of suitable habitats and/or lack of alternative adequate refugia (that could meet their ecological requirements) (Li et al., 2016). The potential role played by evolutionary history characteristics, involved in initially triggering disparate patterns of population genetic structure, is likely to be accentuated by the impact of particular ecobiological features of both species. For instance, it has been postulated that larval dispersal, local retention of larvae as well as local adaptation are the potential processes that could influence population genetic structure across local to intermediate geographic scales (Almany et al., 2013; D’Aloia et al., 2013). As both species were shown to display similar (high) larval dispersal potential (Cuesta and Rodríguez, 2000; Darling et al., 2008; Marino et al., 2011), we hypothesize that other mechanisms including larval retention and ecological requirements (i.e., the potential influence of habitat type) could be potential predictors of levels of gene flow across the African Mediterranean littoral. Nevertheless, while the interplay between impacts of historical and contemporary isolating processes could explain the concordant patterns of mitochondrial and nuclear genetic differentiation recorded within C. aestuarii along the micro- and meso-geographic scales (Deli et al., 2016a), such kind of interaction cannot provide explanation for the mitochondrial and nuclear genetic discordance unraveled in P. marmoratus across the same geographic areas. This could likely stem from the chaotic genetic patchiness found to be characteristic of the population genetic structure of the marbled crab P. marmoratus across micro-geographic scales (Silva et al., 2009; Fratini et al., 2011, 2013; Deli et al., 2016b; Iannucci et al., 2020). At the macro-geographic scale, encompassing the distribution area of both species, although highly significant overall genetic differentiation was retrieved in both species, genetic distances (determined according to nucleotide divergence and haplotype frequency) were remarkably higher in C. aestuarii than the ones recorded in P. marmoratus. This could strongly reflect disparate responses to historical climate fluctuations, but also point out to the impact of temporal extent of genetic diversification. Longer time periods potentially yield much accumulated genetic differences than shorter time spans. This assumption is well in accordance with the determined genetic diversification temporal frames for both species in this study. Trends of historical population dynamics, characterizing C. aestuarii and P. marmoratus, could also play a potential role in shaping spatial distribution of genetic diversity and determining population genetic connectivity. Moreover, preponderant effects of geographic isolation, colonization, migration, and isolation by adaptation are known to be involved in shaping genetic structure at broader scales (Dalongeville et al., 2018).
Lack of significant correlation between pairwise genetic distances determined for Carcinus aestuarii and those assessed in Pachygrapsus marmoratus, from the same locations across the Mediterranean Sea, was recorded at both examined mitochondrial and nuclear genetic markers. This finding suggests that the pattern of pairwise genetic differentiation does not vary in the same way in both examined crab species across the surveyed geographic spectrum of the Mediterranean Sea. Mitochondrial DNA markers have been postulated to be appropriate for inferring species evolutionary histories (Avise, 1994) and assessing the impact of the Quaternary climate oscillations on the distribution dynamics and the build-up of genetic divergence within marine biota (Deli et al., 2018; Zhong et al., 2020). In contrast, biparentally inherited nuclear DNA markers are adequate for inferring fine-scale genetic patterns especially contemporary gene flow mirrored by more recent demographic history (Guo et al., 2021). In light of these considerations, the obtained result not only suggests that both species might have not responded similarly to palaeoclimate conditions but also proposes different kinds of interactions of both decapods populations with contemporary oceanographic and physicochemical characteristics of both Western and Eastern Mediterranean Basins (including temperature and salinity gradients). This assumption could be supported by the outcomes of different isolation mechanisms (IBD, IBE, and IBR) susceptible of driving significant genetic differentiation. Across the defined and selected sites within the Mediterranean Sea harboring both species samples, only C. aestuarii was shown to significantly interact with environment and biogeographical province at the mitochondrial Cox1 gene, and with geography and environment at the nuclear microsatellite loci. The disparate trends of genetic differentiation across the Siculo-Tunisian Strait, resulting in only a significant IBR pattern in C. aestuarii, could reflect the impact of palaeogeographic and palaeoclimate shifts of the Pleistocene on population genetic connectivity of the species across this biogeographic boundary (Deli et al., 2016a). The impact of geography on population genetic structure of the green crab C. aestuarii, discerned only at the nuclear level, while lacking in the marbled crab P. marmoratus, likely suggests the effect of contemporary processes including significant influence of microevolutionary forces, such as genetic drift, on gene pools in the relatively isolated estuarine environments. Provided that both crab species display high larval dispersal potential and fecundity, the significant influence of environment on population genetic structure, discerned only within C. aestuarii, hints at the potential impact of different larval behaviours on generating different patterns of population genetic structure across the Mediterranean. In this context, translocated larvae, corresponding to both species, may be differently adapted to their place of destination leading to a process of isolation by adaptation (Nosil et al., 2009). Numerous investigations have shown that the larvae of marine invertebrates (including crustaceans) are not totally passive, and that, even if they are carried away by currents, the larvae require a suitable site to complete their metamorphosis (Eckman, 1996; Weersing and Toonen, 2009). Hence, larvae of C. aestuarii and P. marmoratus could be exposed to a wide variety of physical, chemical (salinity) and biological (inter- and intra-specific competition, nutrient availability, occurrence of predators) processes that affect the probability of their transition from one stage to another (Wing et al., 1995; Eckman, 1996). This may lead to different patterns of tolerance towards habitat properties including gradual shift in abiotic features such as temperature and salinity. Further experimental investigations on eco-biological properties of the larvae of each decapod species are required to test this scenario.
Temporal frames of genetic diversification were found to be different among Carcinus aestuarii and Pachygrapsus marmoratus based on the analysis of the same aligned fragment length of the mitochondrial Cox1 gene (519 bp). Diversification of the recorded haplotypes within the green crab C. aestuarii started approximately at 0.562 Mya (95% HPD: 0.340-0.801 Mya). For the marbled crab P. marmoratus, genetic diversification time of the examined haplotype dataset dated back to 0.129 Mya (95% HPD: 0.072-0.200 Mya). Notably, for the green crab C. aestuarii, the time frame of genetic diversification, although preceding the ones determined in previous study (Deli et al., 2018), roughly coincides with the glacial period during the MIS Boundary 14/15 at 563000 years before present. This time period nearly corresponds to the late Middle Pleistocene Transition (MPT) between 1.25 Mya and 0.6 Mya (Mudelsee and Schulz, 1997; Clark et al., 2006). During this historical period, the global climate changes were characterized by marked shift in the amplitude and frequency of cold and arid periods, the decreases in sea surface temperatures, as well as the significant increase in ocean levels dropping compared with those of the very Early Pleistocene (Mudelsee and Schulz, 1997; Clark et al., 2006). Accordingly, such palaeoenvironmental and palaeogeographic conditions might have severely affected population genetic structure and dynamics of Mediterranean marine biota including the green crab C. aestuarii. On the other hand, diversification time within P. marmoratus coincides with the Last Interglacial period during the Eemian epoch (between 130000-115000 years ago; Wilson et al., 1998). The Last Interglacial (LIG) was a period characterized by higher global sea level and reduction in ice sheet area (Zagwijn, 1996). It was also marked by warmer climate conditions, which might have been favourable for extended colonization and diversification of marine fauna (Muhs et al., 2002) including the marbled crab P. marmoratus. These two recorded different temporal frames suggest different modes of genetic diversification in both crab species. Glacial vicariance could be at the origin of the onset of the different haplogroups within C. aestuarii as already identified in previous investigations (Deli et al., 2016a, 2018). On the other hand, interglacial dispersal might have been the driving force of genetic diversification within P. marmoratus from penultimate glacial Mediterranean refugia (provided that this region was found to harbor the highest genetic diversity; Deli et al., 2016b; Fratini et al., 2016). In this context, we hypothesize that the recorded trend of mitochondrial genealogy within P. marmoratus could stem from population turnover leading to the erosion of potential genetically divergent lineages, accumulated over previous cyclic glacial maxima (Grant and Bringloe, 2020). Notably, the population-turnover model may provide an explanation for the discerned differences in variation of patterns of population genetic diversity and structure among the two examined decapod species C. aestuarii and P. marmoratus, with quite similar dispersal potential, but with different responses to palaeoenvironmental conditions.
Overall, the present study, using the two coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus as model species, provides relevant insights into the potential origin of disparate patterns of population genetic diversity and structure usually recorded among marine species with quite similar distribution range and larval dispersal potential. It also stresses on the importance of integrating evolutionary history with species ecological requirements and life history traits as well as interaction patterns with geography, environment and biogeographical boundaries in order to unravel the main causes of discordant trends of population genetic polymorphism and structuring across the same surveyed geographic area.
Acknowledgements:
We acknowledge our colleague and co-author, Christoph D. Schubart, who passed away during the build-up to this manuscript, for his important and significant contribution to the study, as well as for all the other valuable and interesting scientific contributions he made during his tragically short but rich life. We also thank two anonymous reviewers for their helpful and constructive comments that contributed to improving the manuscript quality.
Temim Deli, Noureddine Chatti, Khaled Said, Enrique González-Ortegón, Christoph D. Schubart. Population genetic polymorphism and structure in the two coastal crab species Carcinus aestuarii (Brachyura, Carcinidae) and Pachygrapsus marmoratus (Brachyura, Grapsidae), across the Mediterranean Sea, reflect residual effects of different evolutionary histories[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-024-2401-1
Temim Deli, Noureddine Chatti, Khaled Said, Enrique González-Ortegón, Christoph D. Schubart. Population genetic polymorphism and structure in the two coastal crab species Carcinus aestuarii (Brachyura, Carcinidae) and Pachygrapsus marmoratus (Brachyura, Grapsidae), across the Mediterranean Sea, reflect residual effects of different evolutionary histories[J]. Acta Oceanologica Sinica. doi: 10.1007/s13131-024-2401-1
Table
1.
Information on the examined molecular marker, the defined geographic scale, and the selected populations (including the number of samples) for the two coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus (Further details on the geographic origin of the surveyed sampling sites for both decapods species, including countries and basins, are indicated in the references as listed within the Table)
Population
Carcinus aestuarii
Population
Pachygrapsus marmoratus
Cox1 gene
Microsatellite loci
Cox1 gene
Microsatellite loci
Micro-georaphic scale (total number = 110)
Meso-georaphic scale (total number = 133)
Macro-georaphic scale (total number = 412)
Micro-georaphic scale (total number = 90)
Meso-georaphic scale (total number = 110)
Micro-georaphic scale (total number = 60)
Meso-georaphic scale (total number = 80)
Macro-georaphic scale (total number = 587)
Micro-georaphic scale (total number = 90)
Meso-georaphic scale (total number = 110)
Tabarka
10 (a)
10 (a)
10 (a)
10 (a)
10 (a)
Tabarka
02 (d)
02 (d)
02 (d)
10 (d)
10 (d)
Bizerte
15 (a)
15 (a)
15 (a)
10 (a)
10 (a)
Bizerte
06 (d)
06 (d)
06 (d)
10 (d)
10 (d)
Sidi Rais
11 (a)
11 (a)
11 (a)
10 (a)
10 (a)
Korbos
09 (d)
09 (d)
09 (d)
10 (d)
10 (d)
Kelibia
10 (a)
10 (a)
10 (a)
10 (a)
10 (a)
Kelibia
06 (d)
06 (d)
06 (d)
10 (d)
10 (d)
Benikhiar
12 (a)
12 (a)
12 (a)
10 (a)
10 (a)
Benikhiar
06 (d)
06 (d)
06 (d)
10 (d)
10 (d)
Monastir
11 (a)
11 (a)
11 (a)
10 (a)
10 (a)
Monastir
06 (d)
06 (d)
06 (d)
10 (d)
10 (d)
Chebba
12 (a)
12 (a)
12 (a)
10 (a)
10 (a)
Chebba
05 (d)
05 (d)
05 (d)
10 (d)
10 (d)
Sfax
15 (a)
15 (a)
15 (a)
10 (a)
10 (a)
Sfax
09 (d)
09 (d)
09 (d)
10 (d)
10 (d)
Djerba
14 (a)
14 (a)
14 (a)
10 (a)
10 (a)
Zarzis
11 (d)
11 (d)
11 (d)
10 (d)
10 (d)
Tripoli
11 (a)
11 (a)
10 (a)
Tajura
10 (d)
10 (d)
10 (d)
Mosrata
12 (a)
12 (a)
Annaba
10 (d)
10 (d)
Ebro Delta
28 (b)
Terciera
17 (f)
Camargue
20 (b)
Fuerteventura
18 (e)
Livorno
16 (b)
Sesimbra
18 (e)
Elba
16 (b)
Cala Iris
15 (e)
Sardinia
18 (b)
Valencia
20 (e)
Lago Lungo
15 (b)
Tarragona
20 (e)
Fusaro
15 (b)
Saint Florent
15 (e)
Sicily
10 (b)
Calafuria
15 (e)
Venice Lagoon
15 (a, b)
Porto di Follonica
05 (f)
Pomer
20 (b)
Porto Ercole
15 (e)
Amvrakikos
14 (b, c)
Giglio
11 (e)
Lefkada
14 (c)
Rocchette
14 (e)
Navarino
10 (c)
Montecristo
15 (e)
Peloponnesus
06 (b)
Gorgona
09 (f)
Alexandroupolis
12 (c)
Gaeta
15 (e)
Izmir Bay
06 (c)
Fusaro
15 (e)
Enez Dalyan Lagoon
20 (c)
Crotone
16 (e)
Dardanelles Strait
05 (c)
Messina
16 (e)
Prince’s Islands
12 (c)
Porto Cesareo
07 (f)
Bosphorus Strait
07 (c)
Othonoi
07 (f)
Ancona
07 (f)
Otranto
07 (f)
Torre Guaceto
07 (f)
Tremiti
07 (f)
Karaburun Peninsula
07 (f)
Kornati
07 (f)
Pula
18 (f)
Zadar
11 (f)
Boka Kotorska Bay
07 (f)
Crete
14 (f)
Paros Naoussa harbour
07 (f)
Chalkidiki
08 (f)
Beldibi
18 (e)
Girne
22 (f)
Sile
07 (f)
Sinop
07 (f)
Sozopol
21 (f)
Cape Kaliakra
07 (f)
Ropotamo-Kiten
07 (f)
Batumi
07 (f)
Karadag
07 (f)
Tarhankut
07 (f)
Costinesti
07 (f)
Note: (a) Deli et al. (2016a); (b) Ragionieri and Schubart (2013); (c) Deli et al. (2018); (d) Deli et al. (2016b); (e) Fratini et al. (2011); (f) Fratini et al. (2016). The micro-geographic scale corresponds to the Tunisian coast. The meso-geographic scale refers to the North-African coast. It is defined by either the Tunisian and Libyan littorals for the Cox1 data of Carcinus aestuarii or the Tunisian, Algerian and Libyan coasts for the Cox1 data of Pachygrapsus marmoratus. For the examined microsatellite data of both crab species, the meso-geographic scale is represented by the Tunisian and Libyan littorals. The macro-geographic sale corresponds to the distribution range of each examined crab species.
Table
2.
Information on the selected (same or geographically very close) Mediterranean sites harboring the two coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus (examined for mitochondrial and nuclear markers), regions, and the number of samples per location
Selected (same or geographically very close) Mediterranean sites harbouring both species
Region
Cox1 gene
Microsatellite loci
Carcinus aestuarii
Pachygrapsus marmoratus
Carcinus aestuarii
Pachygrapsus marmoratus
Location
Sample number
Location
Sample number
Location
Sample number
Location
Sample number
Site (1)
Western Mediterranean
Ebro
28
Tarragona
20
Site (2)
Western Mediterranean
Livorno
16
Calafuria
15
Site (3)
Western Mediterranean
Elba
16
Porto di Follonica + Rocchette*
19
Site (4)
Western Mediterranean
Lago Lungo
15
Gaeta
15
Site (5)
Western Mediterranean
Fusaro
15
Fusaro
15
Site (6)
Western Mediterranean
Tabarka
10
Tabarka + Annaba*
12
Tabarka
10
Tabarka
10
Site (7)
Western Mediterranean
Bizerte
15
Bizerte
06
Bizerte
10
Bizerte
10
Site (8)
Western Mediterranean
Sidi Rais
11
Korbos
09
Sidi Rais
10
Korbos
10
Site (9)
Western Mediterranean
Kelibia
10
Kelibia
06
Kelibia
10
Kelibia
10
Site (10)
Western Mediterranean
Benikhiar
12
Benikhiar
06
Benikhiar
10
Benikhiar
10
Site (11)
Eastern Mediterranean
Monastir
11
Monastir
06
Monastir
10
Monastir
10
Site (12)
Eastern Mediterranean
Chebba
12
Chebba
05
Chebba
10
Chebba
10
Site (13)
Eastern Mediterranean
Sfax
15
Sfax
09
Sfax
10
Sfax
10
Site (14)
Eastern Mediterranean
Djerba
14
Zarzis
11
Djerba
10
Zarzis
10
Site (15)
Eastern Mediterranean
Tripoli
11
Tajura
10
Tripoli
10
Tajura
10
Site (16)
Eastern Mediterranean
Pomer
20
Pula
18
Site (17)
Eastern Mediterranean
Amvrakikos
14
Othonoi
07
Note: * These geographically very close sites were combined for Pachygrapsus marmoratus in order to increase the sample size.
Table
3.
Comparison of overall genetic differentiation among the two sympatric species Carcinus aestuarii and Pachygrapsus marmoratus across the three defined geographic scales (micro-geographic, meso-geographic, and macro-geographic scales, as indicated in Table 1)
Species/molecular marker examined for genetic differentiation across each geographic scale
Cox1 mitochondrial gene
Microsatellite nuclear loci
Micro-geographic scale (N=60)
Meso-geographic scale (N=80)
Macro-geographic scale (N=412)
Micro-g eographic scale (N=90)
Meso- geographic scale (N=110)
Distance method: Tamura and Nei distance (P)
Distance method: Haplotype frequency (P)
Distance method: Tamura and Nei distance (P)
Distance method: Haplotype frequency (P)
Distance method: Tamura and Nei distance (P)
Distance method: Haplotype frequency (P)
Distance method: number of different alleles (P)
Distance method: number of different alleles (P)
Carcinus aestuarii
ΦST=0.177 (P<0.05)
FST=0.134 (P<0.01)
ΦST=0.233 (P<0.01)
FST=0.116 (P<0.01)
ΦST=0.569 (P<0.001)
FST=0.108 (P<0.001)
FST=0.013 (P<0.01)
FST=0.016 (P<0.001)
Pachygrapsus marmoratus
ΦST=0.043 (P=0.073)
FST=0.028 (P=0.151)
ΦST=0.027 (P=0.130)
FST=0.013 (P=0.263)
ΦST=0.210 (P<0.001)
FST=0.150 (P<0.001)
FST=0.020 (P<0.001)
FST=0.022 (P<0.001)
Note: The computed values for mitochondrial genetic distances were obtained after a rarefaction procedure as described in details in the material and methods section. The values, highlighted for each genetic distance measure, were inferred from the same compared number (N) of samples representing the set of populations included in each defined geographic spectrum (as cited in Table 1). Trend and significance (P) of overall genetic differentiation were calculated from 10000 permutations. Significant values are denoted in bold.
Table
4.
Investigation of the origin of genetic differentiation within sympatric Mediterranean populations of Carcinus aestuarii and Pachygrapsus marmoratus (as indicated in Table 2 and shown in Fig. 1), highlighting patterns of overall genetic differentiation based on the analyses of mitochondrial and nuclear genetic markers and assessing correlations (R) between genetic distances and geographic distances (isolation by distance), environmental distances (isolation by environment), and the distance matrix defined according to the potential influence of the biogeographic boundary at the Siculo-Tunisian Strait (isolation by resistance)
Studied species/Analyzed molecular marker for potential origins of genetic differentiation
Carcinus aestuarii
Pachygrapsus marmoratus
Mitochondrial DNA (Cox1 gene)
Nuclear DNA (microsatellite loci)
Mitochondrial DNA (Cox1 gene)
Nuclear DNA (microsatellite loci)
Implemented genetic distance (overall genetic differentiation and its significance)
Tamura-Nei distance (ΦST=0.221; P=0.000)
number of different alleles (FST=0.016; P=0.000)
Tamura-Nei distance (ΦST=0.011; P=0.223)
number of different alleles (FST=0.024; P=0.000)
Isolation by distance (IBD)
R=0.080 (P=0.352)
R=0.583 (P=0.000)
not applicable
R=0.074 (P=0.628)
Isolation by environment (IBE)
R=0.337 (P=0.000)
R=0.569 (P=0.000)
not applicable
R=−0.157 (P=0.303)
Isolation by resistance (IBR)
R=0.263 (P=0.002)
R=0.219 (P=0.146)
not applicable
R=0.010 (P=0.947)
Note: Significant correlations (P<0.05) are marked in bold. In the case of non-significant overall genetic differentiation, all of the three aforementioned kinds of correlations cannot be computed (not applicable).
Note: (a) Deli et al. (2016a); (b) Ragionieri and Schubart (2013); (c) Deli et al. (2018); (d) Deli et al. (2016b); (e) Fratini et al. (2011); (f) Fratini et al. (2016). The micro-geographic scale corresponds to the Tunisian coast. The meso-geographic scale refers to the North-African coast. It is defined by either the Tunisian and Libyan littorals for the Cox1 data of Carcinus aestuarii or the Tunisian, Algerian and Libyan coasts for the Cox1 data of Pachygrapsus marmoratus. For the examined microsatellite data of both crab species, the meso-geographic scale is represented by the Tunisian and Libyan littorals. The macro-geographic sale corresponds to the distribution range of each examined crab species.
Figure 1. Geographic distribution of the selected seventeen sampling locations harbouring the coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus across the Mediterranean Sea. Details on these localities, including the corresponding number of analyzed samples of each species per site, were highlighted in Table 2. S-T S represents the Siculo-Tunisian Strait. Construction of the base map was carried out by the software DIVA-GIS 7.5.0 (http://www.diva-gis.org).
Figure 2. Variation trends of mitochondrial genetic polymorphism (a. haplotype diversity, b. nucleotide diversity, and c. mean number of nucleotide variation) in the two coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus across the three surveyed geographic scales (as defined in Table 1). For each geographic spectrum, levels of genetic diversity parameters were computed and inferred for the same number of specimens (N) corresponding to both examined species.
Figure 3. Variation trends of nuclear genetic polymorphism (a. expected heterozygosity, b. allelic richness, and c. mean number of alleles per locus) in the two coastal crab species Carcinus aestuarii and Pachygrapsus marmoratus across the two surveyed micro- and meso-geographic scales (as defined in Table 1). For each geographic spectrum, levels of genetic diversity parameters were computed and inferred for the same number of specimens (N) corresponding to both examined species.
Figure 4. Multivariate regression analysis depicting correlation between average genetic dissimilarities (genetic distances) detected among Carcinus aestuarii populations and those discerned among Pachygrapsus marmoratus populations from the same Mediterranean locations (as indicated in Table 2 and exhibited in Fig. 1), for both mitochondrial Cox1 gene (based on Tamura and Nei distances and haplotype frequencies) and nuclear microsatellite loci (based on the number of different alleles).



 DownLoad:
DownLoad:
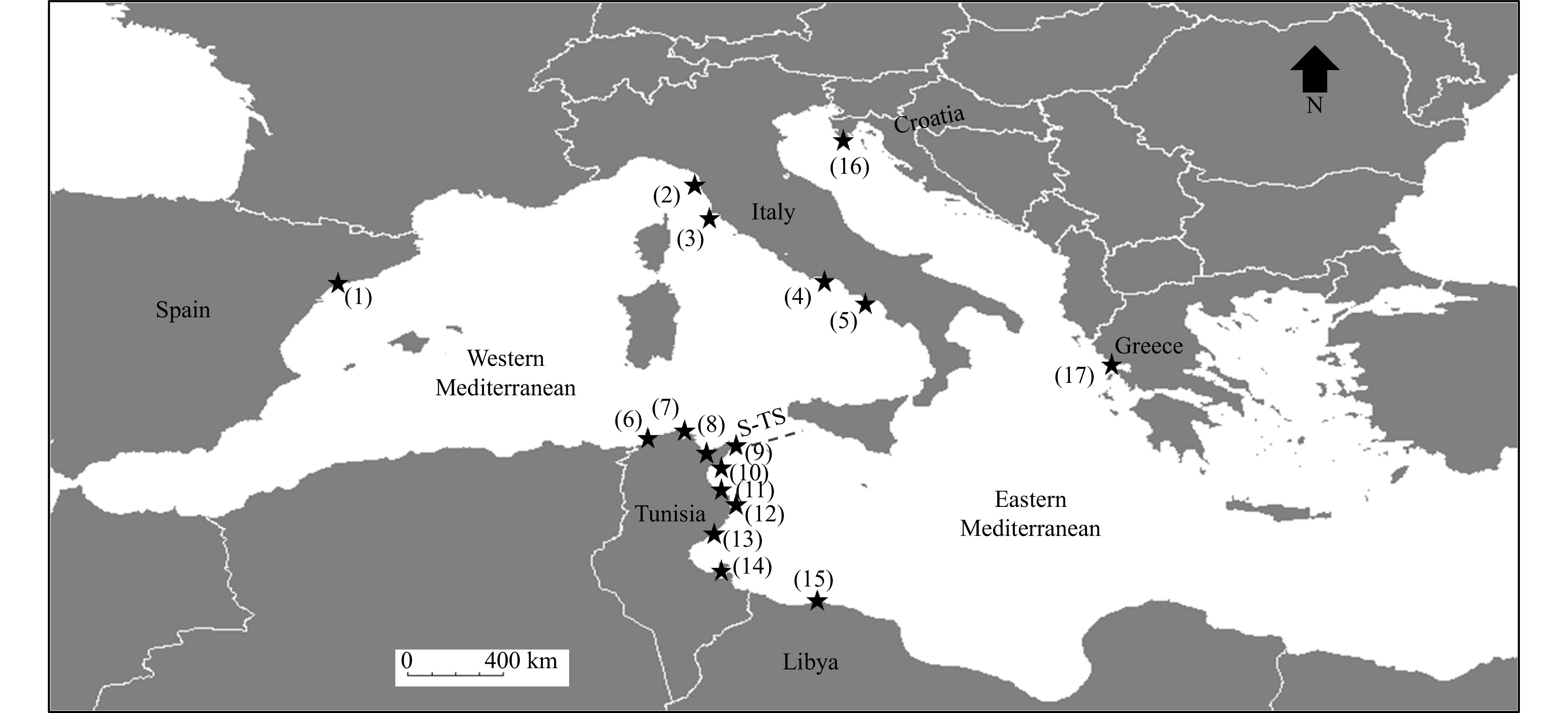
 DownLoad:
DownLoad: