State Key Laboratory of Estuarine and Coastal Research, East China Normal University, Shanghai 200241, China
2.
School of Life Science, East China Normal University, Shanghai 200241, China
3.
School of Life and Environmental Sciences, Hangzhou Normal University, Hangzhou 310018, China
4.
Shanghai Runzhuang Agricultural Science and Technology Co., Ltd., Shanghai 201400, China
Funds:
The Shanghai Chongming Dongtan Bird National Nature Reserve Resource Monitoring Project under contract No. CMDT-JC202101; the Shanghai Municipal Science and Technology Commission under contract No. 19ZR1436900.
An ecological optimization project (semi-closed reclamation project) was implemented to control the invasion of Spartina alterniflora, and optimize the habitat of the Chongming Dongtan wetland, in the Changjiang (Yangtze) River Estuary. After project implementation, a macrobenthic ecological survey was conducted in a natural tidal flat and a semi-closed reclamation restoration area within the Chongming Dongtan wetland from 2019 to 2020. Compared with historical data before reclamation, findings showed that the groups, numbers, and species diversity of the macrobenthos increased significantly, and the ecological optimization project resulted in good ecological benefits. In addition, compared to the natural tidal flat, the number of collected macrobenthic phyla, and the macrobenthic density and biomass were significantly lower in the restoration area. Furthermore, the biodiversity index and functional redundancy of natural tidal flats were generally higher, indicating that the community composition and function of natural tidal flats were relatively more stable. Even though the species composition differed between a number of restoration areas and natural tidal flats, there was no difference in functional diversity, indicating that the effect of restoring ecological functions in restoration areas was optimal. Among them, the biodiversity and functional redundancy of Site S2 were significantly reduced, and the ecosystem function was extremely unstable. Habitat heterogeneity, vegetation community and decreasing salinity were the main factors that affected the ecological functions of macrobenthos. The ecological quality was also evaluated; the Transects N3 and N4 showed good quality. The overall ecological quality of the restoration area was generally high, but that of Site S2 was poor and that of Site S2E was merely good, which was mainly due to modifications of the ecological function of macrobenthos. It is suggested that reeds mowing and freshwater species release should be adopted in restoration areas to improve the community function and the environmental disturbance resistance of the macrobenthos.
Estuaries have some of the highest ecological and economic ecosystem values, and provide a variety of services to human populations (Peterson and Lipcius, 2003; Barbier et al., 2011). However, due to the impact of urban expansion and anthropogenic activities, estuarine ecosystems are facing a series of severe challenges, such as heavy metal pollution (Nunes et al., 2008), sewage discharge (Wang et al., 2021), wetland reclamation (Huang et al., 2021) and biological invasions (Wang et al., 2010). These issues are likely to interfere with the ecological functions and services normally provided by estuaries (Bertness et al., 2002; Laursen et al., 2002; Seitzinger, 2008). The Changjiang (Yangtze) River Estuary comprises the largest estuarine wetland resources in China (Nie et al., 2015), however, in its wetland areas, Spartina alterniflora invasion has been a major source of disturbance over a long period. In the Dongtan wetland of Chongming Island, such invasion has caused negative effects on other saltmarsh vegetation, waterfowl habitats and macrobenthic communities (Ma et al., 2017). To control the spread of this exotic species and restore bird habitats, a project was established by the State Forestry Administration of China and Shanghai City Government in 2011, officially known as “the ecological control of S. alterniflora and the optimization of bird habitats in the Chongming Dongtan wetland” (Tang, 2016). This ecological optimization project is also known as a semi-enclosed reclamation restoration project. The water level was increased through embankment constructions and sluice control. Thereby, S. alterniflora was reaped under water and its residual roots died due to oxygen deprivation. Giving that reclamation has a negative impact on the original habitat (Lv et al., 2016a), a long term and systematic evaluation of the restoration area needs to be conducted after the ecological optimization project in the Chongming Dongtan wetland.
As an important part of estuarine wetland ecosystems, the macrobenthos maintains key ecological functions, including energy exchange and nutrient circulation within the food web (Herman et al., 1999; Borja et al., 2012). Modifications of these communities affect both the upper and lower trophic levels, represented by organisms such as birds and algae, respectively (Sakamaki and Richardson, 2009; Christianen et al., 2017). Moreover, microbenthic organisms are very vulnerable to the direct stress posed by changes in the estuarine ecosystem due to their sedentary and attached life mode. Therefore, it is suggested that macrobenthic community functions and ecological assessment should be regarded as important biological indicators to monitor and evaluate changes in estuarine ecosystems (van der Linden et al., 2012; Piló et al., 2016). By monitoring the dynamic changes of the macrobenthic community, the ecological effects before and after reclamation of the Changjiang Estuary wetland can be studied, and the ecological quality of the benthic habitat after reclamation can be evaluated.
Species diversity is closely related to the biogeochemical functions of estuaries, such as productivity, blue carbon storage, and nutrient cycling (Witman et al., 2008; Mascaro et al., 2012; Gao and Lin, 2018). However, traditional metrics such as species richness, abundance, diversity, and functional groups ignore the similarities and differences between interspecific functions, providing only limited information about the ecological functions of such communities (Poos et al., 2009; Schmera et al., 2009; van der Linden et al., 2012). Therefore, functional diversity is often introduced when evaluating the ecological functions of the macrobenthic communities to describe their functional characteristics. Rao’s quadratic entropy coefficient and the functional dispersion index are the two main indices commonly used (Han et al., 2021).
The relationship between diversity and ecosystem functions is based on the distribution pattern of species in ecological niches or functional spaces. Functional niche complementarity and overlap have been suggested as a possible mechanism linking diversity to ecosystem processes (Tilman et al., 2001; Adler and Bradford, 2002; Petchey, 2003). In other words, several species play a similar role regarding their ecological functions; thus, they do not necessarily change due to the loss or replacement of species (Petchey and Gaston, 2002, 2006; Rosenfeld, 2002). Therefore, the concept of functional redundancy is introduced to test the stability of ecosystem functions after the disappearance of some species (Petchey and Gaston, 2006). Regarding the characteristics of ecological functions associated with the macrobenthos, previous studies have mainly focused on mangroves (Leung, 2015), freshwater river systems (Voß and Schäfer, 2017; Chen et al., 2018), and some estuarine systems (van der Linden et al., 2012; Piló et al., 2016).
An index related to the macrobenthos is also suitable for the assessment of estuarine ecosystem health. Presently, dozens of methods have been developed to assess the health of aquatic environments through the analysis of macrobenthic communities. Among them, abundance-biomass comparison curves (ABC curves) are also commonly used to monitor the effects of human disturbance or pollution on macrobenthic communities (Warwick, 1986; Liu et al., 2018a). Out of the several biotic indices on benthic macroinvertebrates available globally (Borja et al., 2000, 2015), the AZTI’s Marine Biotic Index (AMBI) and the multivariate AZTI’s Marine Biotic Index (M-AMBI) (Muxika et al., 2007) were selected in the present study. The metrics and biotic indices mentioned above have been increasingly used in recent years to assess the ecological quality status (EQS) of estuaries (Luo et al., 2016; Zhou et al., 2018; Huang et al., 2021). Nevertheless, the use of a single biological index to assess ecological quality may be too partial due to the complexity of these ecosystems. Therefore, multiple appropriate biological metrics and indices should be integrated to obtain a more comprehensive assessment of the EQS of estuaries (Salas et al., 2006; Martinez-Haro et al., 2015; Liu et al., 2019).
In this study, after the implementation of the ecological optimization project (a semi-closed reclamation project) to remove S. alterniflora, the macrobenthic community structure and ecological health of the reclamation restoration area and a natural tidal flat were systematically studied in the Chongming Dongtan wetland in the Changjiang (Yangtze) Estuary. The main research objectives were: (1) to explore the impact of the reclamation project on the macrobenthic community in the reclamation restoration area and natural tidal flat in Chongming Dongtan wetland by comparing findings with historical data before reclamation; (2) to analyze the effects of different benthic habitats on macrobenthic biodiversity and functional redundancy after reclamation; and (3) to compare and evaluate the ecological quality of the reclamation restoration area and natural tidal flat in the Chongming Dongtan wetland after project implementation.
2.
Materials and methods
2.1
Study area
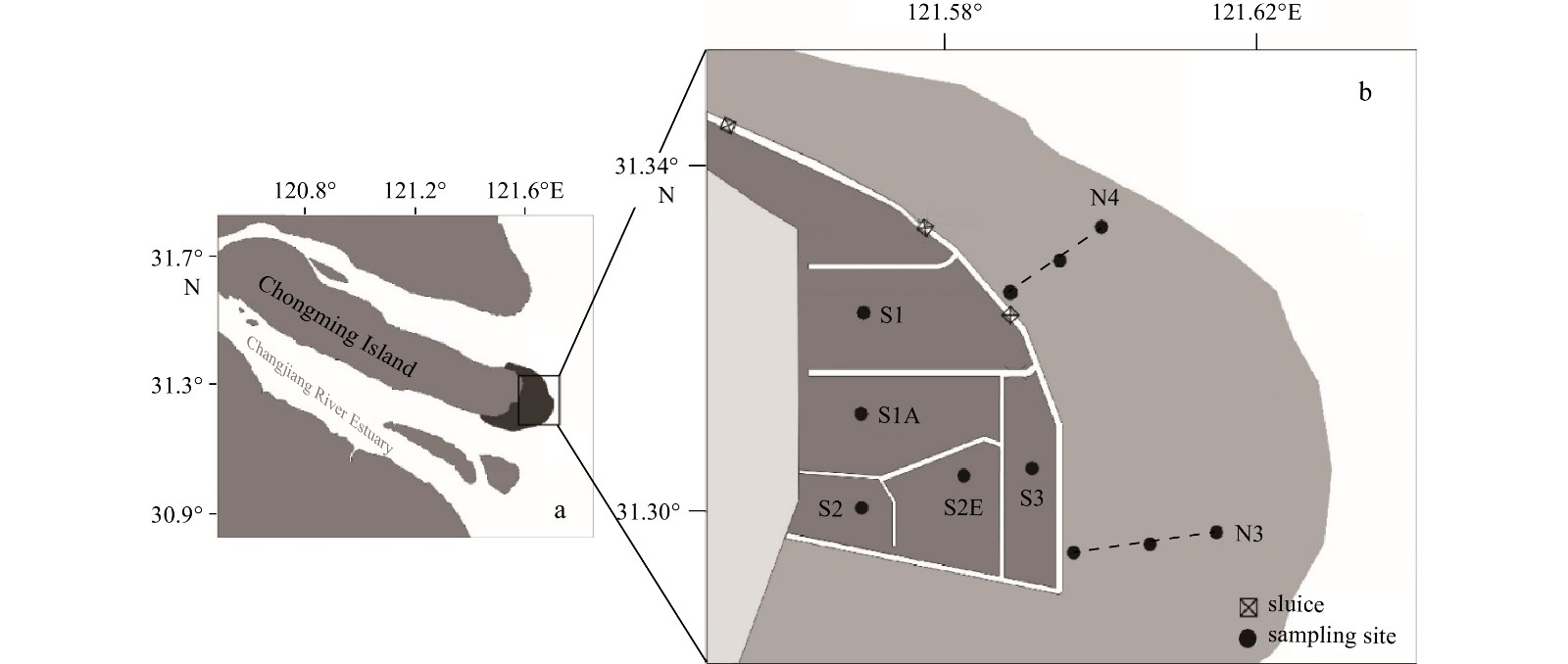
The Chongming Dongtan wetland (31°25'−31°38'N, 121°50'−122°05'E), located in the easternmost part of Chongming Island, is the largest and most well-developed estuary tidal-flat wetland in the Changjiang River Estuary, and is one of the most ecologically sensitive areas in the world (Fig. 1). This wetland represents the breeding ground for several aquatic species that thrive in this area (Forde et al., 2015), and is also a transit station and overwintering location for migratory birds on the migration route from East Asia to Australia (Gao and Zhang, 2006). Due to its important role in maintaining biodiversity and valuable resources, the area was listed as an internationally important wetland by the Ramsar International Convention in 2002 and as a national nature reserve in 2005 to protect its ecological integrity (Tian et al., 2008).
Figure
1.
Schematic diagram of macrobenthic sampling in Chongming Dongtan wetland from 2019 to 2020. In b, the dark grey area represents the semi-closed reclamation restoration area, and the light grey area represents the natural tidal-flat area. The dots represent the sampling sites; the dash lines represent the Transects N3 and N4.
To control the expansion of invasive species, initiatives such as the ecological control of S. alterniflora and bird habitat optimization were carried out in the area. In 2012, a new dike was built in the northern area of the wetland, which included three sluice gates connecting the internal and external water systems. The gates are opened regularly to maintain the water level in the restoration area, by mixing the inner fresh water and outer seawater flows. During the closing period, the water level in the restoration area increases, while during the opening period, it decreases. As a semi-closed reclamation project, it encloses the high and middle tidal flats of the original wetland within the embankment. In the early stage of the project, S. alterniflora was harvested, while reed communities were maintained. The increasing water level in the area flooded S. alterniflora residues and contributed to their complete eradication. There are two types of habitats in the study area: (1) a natural tidal flat area, the low tidal flat area reserved outside the project; and (2) a semi-closed reclamation restoration area.
2.2
Sampling design
In both the natural tidal-flat area and semi-closed reclamation restoration area, sampling surveys were conducted seasonally in May (spring), July (summer), September (autumn), and December (winter) in 2019; and in June (spring), August (summer), October (autumn), and December (winter) in 2020. Two transects (N3, N4) were set in the natural tidal-flat area and five sampling sites (S1, S1A, S2, S2E and S3) were set in the restoration area, based on the habitat characteristics of the Chongming Dongtan Nature Reserve (Fig. 1). In the natural tidal-flat area, along the Transect N3, the vegetation comprised reeds communities and Scirpus mariqueter. Here, the vegetation is divided into blocks by winding tidal creeks. Transect N4 was sparsely vegetated and consisted mainly of dams, rocks, and gravel. In the restoration area: in Site S1 vegetation comprised Hydrilla verticillata and plot-like reeds, growing at a water depth of approximately 35 cm; in Site S1A, the land vegetation was characterized by Phragmites australis, and here the river water level exceeded 1 m all year round; Site S2 had a lot of tall and dense reeds, and was greatly affected by human activities; and sampling site S2E was covered by dense reeds and weeds, and the river water level exceeded 1 m all year round. Scirpus mariqueter was present in Site S3, growing at a water depth of approximately 15 cm.
Three sampling levels were set in the natural tidal-flat area: 50 m, 500 m and 1 000 m offshore; and two sampling levels were set in the restoration area: vegetation absent and vegetation present. Quantitative biological samples (25 cm×25 cm×30 cm) of four biological replicates were collected from the selected sampling areas, and the macrobenthos was separated using a sieve with a 0.45 mm mesh (Huang et al., 2021). At the same time, based on the organisms present, qualitative biological samples were obtained as much as possible. Biological samples were preserved with EtOH 75%, and organisms were identified as accurately as possible to species level. After identification, the specimens were weighed on a scale with an accuracy of 0.001 g. During surveys, a hand-held GPS locator (Chengdu Wanggan Co. Ltd., China) was used for the exact localization of each sampling site. Environmental parameters such as water temperature, salinity, pH, and dissolved oxygen (DO) concentration were measured in situ using a HQ 40D portable multi-parameter water quality analyzer (HACH, USA).
2.3
Biodiversity and function redundancy
Species and taxonomic diversity indices and functional diversity indices, were selected to comprehensively characterize benthic macroinvertebrate communities and evaluate their ecological functions in different habitats within the Chongming Dongtan wetland. The multivariate statistical software Primer v5.0 (PRIMER-E Ltd., New Zealand) was used to calculate the species and taxonomic diversity indices at the species level, while the functional diversity index was calculated using the FD software package in R v4.0.4 (the R Foundation, USA). See Text S1−S3 for more information on the indices.
2.3.1
Species diversity
Based on the quantitative data obtained from species, the Margalef species richness index (d) (Margalef, 1969), Pielou species evenness index (J′) (Levins, 1970) and Shannon-Wiener index (H') (Shannon and Weaver, 1962) were calculated to evaluate species diversity.
2.3.2
Taxonomic diversity
Average taxonomic distinctness (Δ+) and variation in taxonomic distinctness (Λ+) were selected to evaluate the taxonomic diversity of the macrobenthic communities. Before the calculations, the macrobenthos data recorded in the Changjiang River estuarine region between 2010 and 2020 were compiled into a complete list of all the detected species, classified according to the standard six taxonomic grades: Phylum, Class, Order, Family, Genus and Species (Table S1). The two above-mentioned taxonomic diversity indices were calculated based on the species’ qualitative data.
2.3.3
Functional diversity
Twelve functional traits were selected to evaluate functional diversity, namely (1) mobility, (2) life span, (3) body size, (4) adult habitat, (5) reproduction types, (6) reproduction frequency, (7) larval type, (8) larval development, (9) feeding habits, (10) salinity, (11) substrate, and (12) AMBI ecological group (Table S2) (Usseglio-Polatera et al., 2000; Piló et al., 2016; Liu et al., 2018a). Rao’s quadratic entropy (FDQ) and functional dispersion (FDis) values were calculated in combination with relative species abundance. For this parameter, a lg (x+1) conversion was performed to reduce the influence of the dominant species.
2.3.4
Functional redundancy
The ratio between the functional diversity index (FDQ) and H' was used to measure functional redundancy (FDQ/H'). The higher the ratio value, the lower the functional redundancy (van der Linden et al., 2012), indicating the loss of functional niches, resulted in the change and loss of ecological functions. In contrast, a high functional redundancy value means that there are different species in the community performing similar or equal ecological functions, thus the loss of one species has a more limited impact on the overall ecosystem functions (Petchey and Gaston, 2006).
2.4
Analysis of community structure
Community hierarchical clustering and non-metric multidimensional scaling (NMDS) were used to describe community similarities among the macrobenthos in each study area. The analysis was performed in SPSS v23.0 (IBM Corp., USA).
2.5
EQS
2.5.1
ABC method
ABC curves were used to evaluate the disturbance in the macrobenthic communities by comparing the cumulative dominance curves of species density and biomass (Lv et al., 2016b; Ren et al., 2016; Yang et al., 2017). There are three possible scenarios described by this method: (1) the habitat is not disturbed or is slightly disturbed (the biomass curve is located above the abundance curve, indicating that the community structure is in a state of equilibrium); (2) the habitat is moderately disturbed (the biomass curve crosses or overlaps the abundance curve, indicating that the community is disturbed due to pollution-induced stress); and (3) the habitat is severely disturbed (the biomass curve is below the abundance curve).
Warwick and Clarke (1994) proposed the application of W value statistics to ABC curves, and the calculation formula is shown in Text S4.The range of the W value is [−1, 1]. When it approaches 1, the environment is not disturbed or slightly disturbed and the community biomass is gradually dominated by a single species; however, the overall species richness tends to be the same. When the W value tends towards 0, the environment is slightly disturbed or moderately disturbed, and the number of large and competitive dominant species decreases. Finally, when the W value gradually approaches −1, the environment is severely disturbed and species with a small body size become dominant. The ABC method was performed using Primer v5.0.
2.5.2
AMBI and M-AMBI
In the AMBI, macrobenthic organisms are divided into five ecological groups according to their levels of sensitivity to environment disturbance (Borja et al., 2000). Thus, species were scored I to V, and those that were not included in the list were replaced by other species of the same genus or family (Borja et al., 2008). The correspondence between the AMBI value and the EQS is shown in Table 1.
Table
1.
The relationship between AMBI and M-AMBI and ecological quality status/environmental disturbance
Ecological quality/Disturbance classification
AMBI
M-AMBI
High/undisturbed
(0, 1.2]
>0.77
Good/slightly disturbed
(1.2, 3.3]
(0.53, 0.77]
Moderate/moderately disturbed
(3.3, 5.0]
(0.38, 0.53]
Poor/heavily disturbed
(5.0, 6.0]
(0.20, 0.38]
Bad/extremely disturbed
>6.0
(0, 0.20]
Note: AMBI is the abbreviation of AZTI’s Marine Biotic Index; and M-AMBI, multivariate AZTI’s Marine Biotic Index.
The M-AMBI integrates H', species richness, and the AMBI into a factor analysis were used to obtain the pollution status of the eco-environment (Muxika et al., 2007). The relationship between the M-AMBI and EQS is shown in Table 1. AMBI and M-AMBI were calculated using the software AMBI v6.0 (which is freely available at http://ambi.azti.es) and an updated species list.
2.6
Data analysis
SPSS v23.0 statistical software was used to conduct a two-way ANOVA (sample sites and seasons) for the density, biomass, and multiple biodiversity indices, and the Tukey honestly significant difference (HSD) test was used to compare the differences among multiple ecological indicators (P<0.05). Spearman correlation analysis was performed for multiple biodiversity indices using R v4.0.4, and correlation coefficient graphs were drawn using “corrplot” libraries. The histogram was generated in GraphPad Prism v8.0.2 (GraphPad Software, USA).
3.
Results
3.1
Environmental factors
There was no significant difference between dissolved oxygen, pH, and water temperature between the natural tidal flat and the restoration area. The salinity of the latter was significantly lower. Among sample sites in the restoration area, the salinity of Site S3 was the highest, while it was the lowest in Site S2, where the water desalination phenomenon was the most obvious (Table 2).
Table
2.
The biodiversity index of macrobenthos in the Chongming Dongtan wetland, Shanghai from 2019 to 2020
Transect N3
Transect N4
Site S1
Site S1A
Site S2
Site S2E
Site S3
DO concentration/(mg·L−1)
7.95±2.72
7.89±4.54
8.40±3.50
10.09±4.92
11.45±3.84
8.63 ± 2.69
10.49 ± 2.61
pH
8.14±0.47
7.88±0.52
8.02±0.49
8.32±0.29
8.36±0.49
7.96±0.75
8.09±0.50
Water temperature/°C
20.72±10.11
20.30±10.04
19.73±7.77
19.82±8.41
20.49±8.13
19.71±8.38
20.52±8.64
Salinity
8.33±1.91b
8.77±2.27b
3.52±2.65a
3.46±2.67a
1.37±2.07a
3.11±1.97a
4.37±1.96a
d
2.18±0.59c
1.85±0.59bc
0.94±0.42ab
1.67±0.87bc
0.19±0.26a
1.44±0.60bc
1.23±0.74b
J'
0.62±0.15ab
0.65±0.14b
0.70±0.15b
0.82±0.10b
0.18±0.29a
0.60±0.19ab
0.77±0.15b
H'
2.36±0.65b
2.31±0.59b
1.62±0.39b
2.54±0.81b
0.31±0.59a
1.86±0.72b
2.01±1.00b
Δ+
88.33±3.73b
83.43±14.02b
84.74±9.01b
82.47±9.55b
48.26±51.67a
85.24±2.70b
90.13±6.84b
$\varLambda $+
437.13±108.99b
362.08±65.10ab
331.40±166.21ab
533.50±414.53b
51.12±121.58a
521.72±113.81b
332.89±234.43ab
FDQ
42.56±3.20b
44.97±4.10b
37.81±7.68b
41.88±6.15b
11.99±13.81a
38.92±6.67b
39.23±11.46b
FDis
6.42±0.25b
6.60±0.33b
6.01±0.69b
6.39±0.49b
2.20±2.47a
6.15±0.55b
6.09±1.09b
FDQ/H'
19.38±6.34
20.86±7.37
24.49±7.44
17.89±5.95
39.50±53.30
25.03±12.74
24.54±14.46
Note: d is the Margalef species richness index; J', Pielou species evenness index; H', Shannon-Wiener index; Δ+, average taxonomic distinctness; $\varLambda $+, variation in taxonomic distinctness; FDQ, functional diversity index; FDis, functional dispersion; and FDQ/H', functional redundancy. Different letters indicate significant differences among sites.
3.2
Species composition, abundance and biomass of the macrobenthos
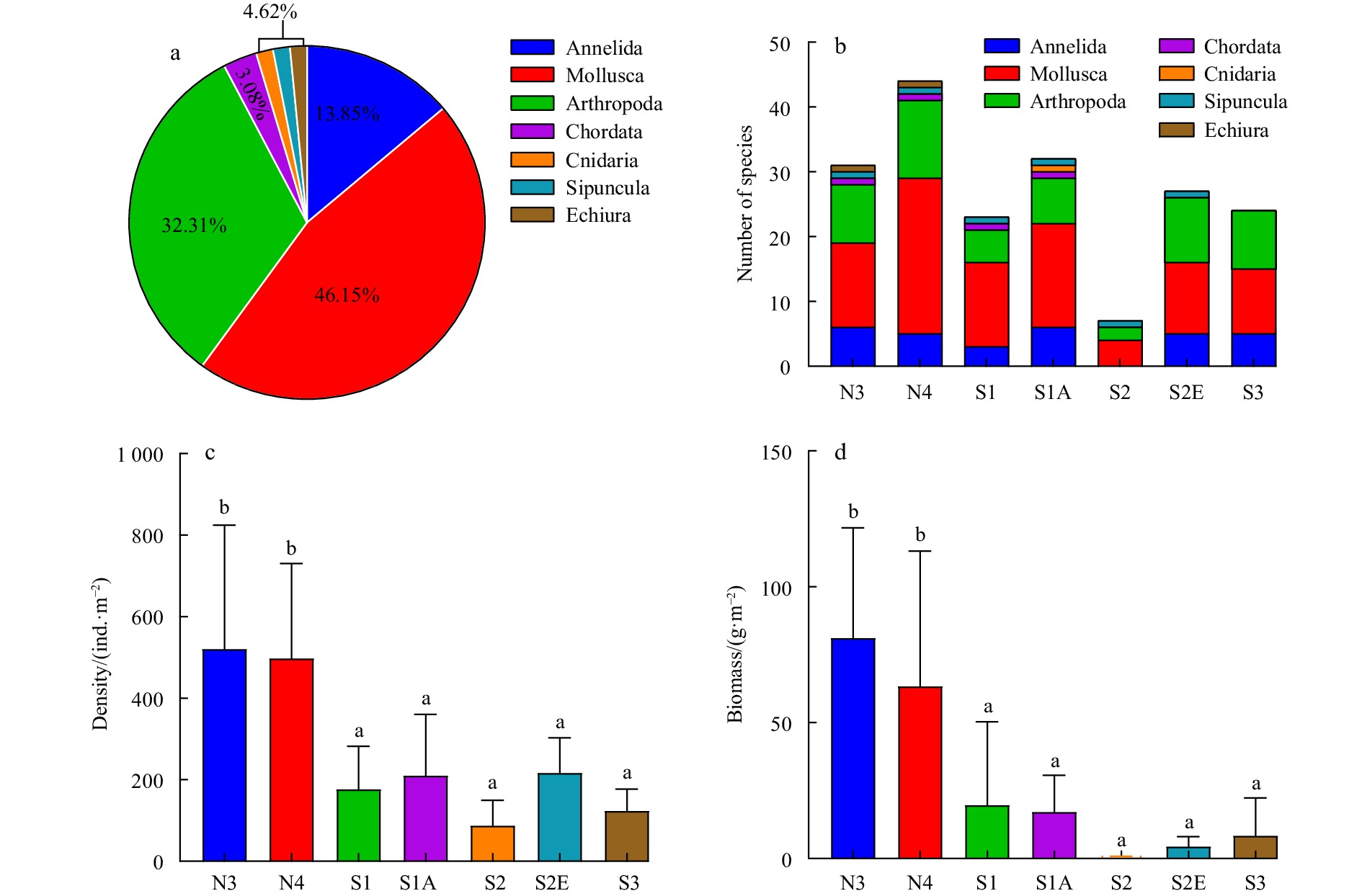
In total, 65 species, 46 families and 7 phyla of macrobenthos were collected from the Chongming Dongtan wetland areas (Fig. 2a). In particular, 30 Mollusca (46.15%), 21 Arthropoda (32.31%), 9 Annelida (13.85%), 2 Chordata (3.08%), and 3 other species (4.62%), belonging to Cnidaria, Sipuncula, and Echiura orders, respectively.
Figure
2.
The species composition (a), number of species (b), density (c) and biomass (d) of macrobenthos in Chongming Dongtan wetland from 2019 to 2020. N3 and N4 are sampling transects; S1, S1A, S2, S2E and S3 are sampling sites. Different letters above the bars indicate significant differences among sites.
In the natural tidal flat, the number of species ranged from 31 to 44. In the restoration area, the number of phyla and species was slightly lower; the number of species ranged from 23 to 32 (except for Site S2), and several of the marine species of mollusks, arthropods, and echiura among them were not found in the restoration area. Seven species were detected in Site S2, consisting only of four mollusks, two arthropods and one sipunculan (Fig. 2b). For the detailed species composition of the sampled wetland areas, see Table S3 of the supporting information.
Compared with the natural tidal flat area (Transects N3 and N4), the density and biomass of the macrobenthos in five sample sites in the restoration area were significantly lower (two-way ANOVA, P<0.05). The density and biomass of the macrobenthos in Site S1 were 57.7% and 67.7% lower, respectively, while for Site S1A, these parameters were 54.2% and 70.9% lower, respectively; they were the lowest in Site S2, specifically 73.4% and 98.7% lower than the values observed in natural tidal flat, respectively. In Site S2E, the density and biomass were 50.7% and 92.2% lower, respectively, and in Site S3, they were 69.0% and 89.9% lower, also respectively (Figs 2c, d). Tables S4 and S5 show the dominant species of each sample site and the types of biological traits they represent.
3.3
Biodiversity analysis
Compared with the other six sampling points, the species diversity (d, J', H'), taxonomic diversity (Δ+, Λ+), and functional diversity (FDQ, FDis) of Site S2 were significantly lower (two-way ANOVA, P<0.05), by 83.2%, 66.0%, and 69.4%, respectively. The species richness index (d) of Sites S1 and S3 in the restoration area was significantly lower (by 54.4% and 41.2%, respectively) than that of Transect N3 in the natural tidal flat (two-way ANOVA, P<0.05). In addition, there was no significant difference in functional redundancy among the above-mentioned sample sites, but Site S2 had the largest value of FDQ/H' and the lowest functional redundancy value (Table 2).
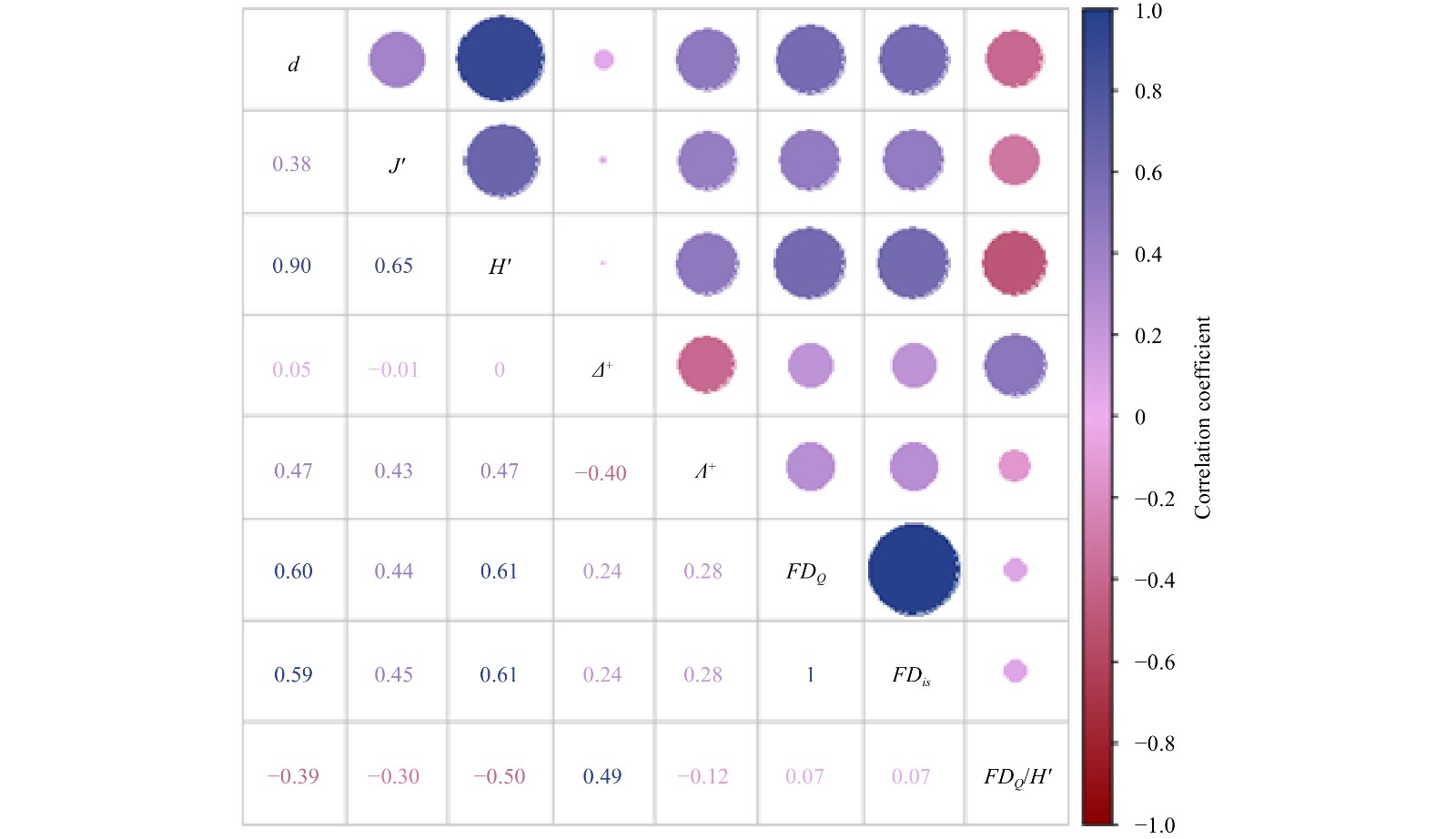
Spearman analysis (Fig. 3) showed that species diversity (d, J', H') was significantly and positively correlated with the index of variation in taxonomic distinctness (Λ+, 0.43−0.47), Rao’s quadratic entropy (FDQ, 0.44−0.61), and the functional dispersion index (FDis, 0.45−0.61) (P<0.01); the taxonomic differential variation index (Λ+) showed the same type of correlation with Rao’s quadratic entropy (FDQ, 0.28) and with the functional dispersion index (FDis, 0.28) (P<0.05); while functional redundancy (FDQ/H') was significantly and negatively correlated with species richness (d, −0.39). The Shannon-Wiener diversity index (H', −0.5) (P<0.01), was also significantly and negatively correlated with species evenness (J', −0.3) (P<0.05), but significantly and positively correlated with the taxonomic differential variation index (Δ+, 0.49) (P<0.01).
Figure
3.
The correlation matrix of the macrobenthic diversity index in Chongming Dongtan wetland from 2019 to 2020. The d is the Margalef species richness index; J′, Pielou species evenness index; H', Shannon-Wiener index; Δ+, average taxonomic distinctness; Λ+, variation in taxonomic distinctness; FDQ, functional diversity index; FDis, functional dispersion; and FDQ/H', functional redundancy. Species diversity includes d, J' and H'; taxonomic diversity, Δ+ and Λ+; functional diversity, FDQ and FDis; Functional redundancy, FD/H'.
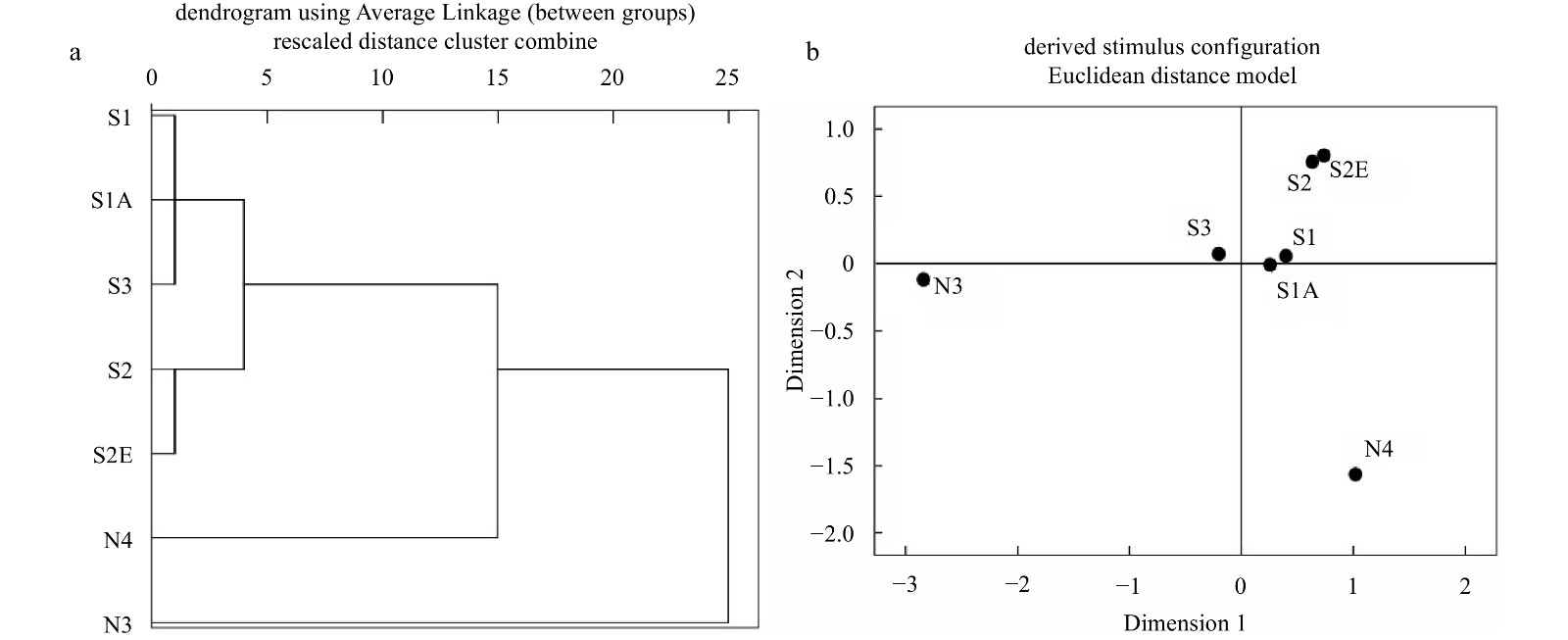
The seven sampling areas were divided into four groups visible in the community cluster tree map (Fig. 4a): (1) Sites S1, S1A and S3; (2) Sites S2 and S2E; (3) Transect N4 and (4) Transect N3. The NMDS results showed that the stress was 0.134, and the distance further confirmed the tree map results (Fig. 4b).
Figure
4.
Clustering and non-metric multidimensional scaling (NMDS) analysis of macrobenthic communities in Chongming Dongtan wetland from 2019 to 2020.
As observed in the cumulative dominance curves of abundance and biomass (Fig. 5), the abundance and biomass of Transect N4 in the natural tidal flat and those of Site S2E in the restoration area overlapped or crossed each other, and the W value approached 0. The ecological health status of Sites N4 and S2E was slightly disturbed (Figs 5b, f). However, the cumulative dominance curves of Site S2 in the restoration area intersected multiple times, and the W value was smaller than 0. The ecological health of Site S2 was moderately disturbed (Fig. 5e). In the remaining habitats, the biomass curve was above the abundance curve, and their ecological health status was not disturbed with the W value close to 1.
Figure
5.
Abundance-biomass comparison curves of 7 habitats in Chongming Dongtan Wetland from 2019 to 2020 (green triangle is abundance; blue triangle, biomass).
Based on the AMBI species list and calculations of species allocation and substitution, Periophthalmus cantonensis was ignored in Transect N4 due to its low abundance and it was also not included in the species list. The AMBI values of the seven sampling points were between 1.453 and 3.014 (which were in the 1.2−3.3 default range), indicating that their ecological health status was slightly disturbed. The M-AMBI values showed that the ecological health status of Site S2 in the restoration area was of poor quality, while that of Site S2E was of good quality. The remaining habitats could be classified as high quality. The specific results are shown in Table 3.
Table
3. AMBI and M-AMBI of 7 habitats in Chongming Dongtan wetland from 2019 to 2020
Sampling area
AMBI
Disturbance classification
M-AMBI
Ecological quality
N3
2.210
slightly disturbed
0.83
high
N4
1.514
slightly disturbed
0.95
high
S1
1.474
slightly disturbed
0.82
high
S1A
1.453
slightly disturbed
0.99
high
S2
2.863
slightly disturbed
0.35
poor
S2E
3.014
slightly disturbed
0.69
good
S3
2.543
slightly disturbed
0.77
high
Note: AMBI is the abbreviation of AZTI’s Marine Biotic Index; and M-AMBI, multivariate AZTI’s Marine Biotic Index.
4.1
Effects of reclamation project on macrobenthos community after S. alterniflora Removal
The community succession of the macrobenthos can reflect changes in benthic habitats in wetlands (Nishijima et al., 2014). According to the research data collected before the embankment construction in 2011 (Chen et al., 2009), there were 15 species in the high and middle tidal flats, and 19 species in the low tidal flats in the original Chongming wetland (Table 4). Conversely, the number of species in the reclamation restoration area and the natural tidal flat increased to 23 and 33, respectively, and the species composition was also richer in this study. Before reclamation, the dominant species in the high and middle tidal flats was Assiminea violacea, while in the low tidal flats, Assiminea violacea, Elachisina sp., and Glaucomya chinensis were the most dominant. After eight years of the optimization project, the number of dominant species in the restoration area and the natural tidal flats increased from 9 to 13, respectively. Among them, the new dominant species in the restoration area after reclamation were mostly euryhaline species, and Assiminea violacea, the original dominant species, no longer had an ecological advantage in the area. The average density of the macrobenthos in the restoration area and natural tidal flat was also significantly higher than that before reclamation in 2011. This could be explained by the removal of S. alterniflora in the semi-closed restoration project, which had a specific positive effect on macrobenthos recovery. In this study, more species were collected in the natural tidal flat than in the restoration area, their species composition was relatively richer, and their density and biomass were also significantly higher. In addition, in the restoration area, larger mollusks (such as oysters, and Glauconome chinensis) and arthropods (such as Chiromantes dehaani, Gelasimus arouatus, etc.) were hardly found, while large- and medium-sized mollusks and arthropods were distributed across the natural tidal flat. This indicated that the natural tidal flats after reclamation were more conducive to macrobenthos survival and reproduction. Ecological restoration projects focus on evaluating and protecting biodiversity, and species diversity is a common indicator of restoration. When comparing the macrobenthic species diversity before and after reclamation, the species diversity indices (d, J', H') increased significantly after reclamation. Compared with the data before and after reclamation, the ecological optimization project optimized the new habitat after the removal of S. alterniflora, resulting in a significant increase in the number of species, density, and diversity of the macrobenthos, as well as a gradual increase in the number of dominant species and a richer species composition. The results showed that the removal of S. alterniflora by the ecological optimization project had a positive effect on macrobenthos, and that the macrobenthic community recovered significantly. The ecological optimization project (semi-closed reclamation project) can represent an ecologically effective approach to control S. alterniflora invasion, which can result in positive ecological effects in the wetland environment and biological community.
Table
4. Comparison of community characteristics before and after reclamation in Chongming Dongtan wetland
Variable
2007−2008
2019−2020
High and middle marsh
Low marsh
Semi-closed restoration area
Natural tidal flat area
Average number of species
15
19
23
33
Average density/(ind.·m−2)
672.75
1866.5
1304.8
4078.5
d
2.054
2.461
3.063
3.851
J'
0.514
0.584
0.587
0.634
H'
1.946
2.502
2.707
3.195
Note: d is the Margalef species richness index; J', Pielou species evenness index; H', Shannon-Wiener index.
4.2
Effects of different benthic habitats on biodiversity and functional redundancy of macrobenthos after reclamation
The key to ecological protection and restoration is to maintain the health and stability of ecosystem functions (Thorpe and Stanley, 2011). Some studies have also pointed out that the comprehensive analysis and evaluation of biodiversity using multiple indices can reveal different types of information (Korb et al., 2016; Li et al., 2019). Except for Site S2, the species richness index in several locations in the restoration area was lower than that in the natural tidal flat area. However, the species evenness index and the Shannon-Wiener diversity index in the restoration area were not significantly different from those in the natural tidal flat. There was a positive correlation between the variation index and species diversity, while theoretically, the mean variation index was not strongly correlated with species diversity (Zhu et al., 2018). This indicates that there were differences in terms of species diversity between the restoration area and the natural tidal flat, but the phylogenetic relationship between them and the uniformity of the species classification were basically the same. Therefore, the macrobenthic diversity of the restoration area was only slightly different from that of the natural tidal flat area. Functional diversity provides a new approach to the study of ecosystem processes and ecological functions (Petchey and Gaston, 2006; van der Linden et al., 2012). In the present study, the results showed that functional diversity and functional dispersion were more closely related to species diversity. Although species richness decreased in the restoration area, there was no difference in functional diversity and dispersion between the restoration area and the natural tidal flat (except for Site S2). A high FDis value can indicate that the distribution of species traits is relatively uniform, functional characteristics are relatively consistent, and all species have similar weighted distances from the mass center. Studies have shown that communities with high functional differentiation have a higher efficiency in the utilization of resources (Zhong et al., 2020). In the present study, the resource utilization efficiency of the macrobenthos in the natural tidal flat was similar to that in the restoration area, even though the species composition differed. In addition, the species richness, density, biomass, and biodiversity of Site S2 in the restoration area clearly decreased, and the community structure in this area changed significantly.
Most studies have also found that the functional diversity of the macrobenthos is positively correlated with habitat heterogeneity (Wong and Dowd, 2015; Liu et al., 2018b). Within the natural tidal flat, Transect N3 consisted of a complex habitat of interwoven tidal creeks characterized by the presence of Scirpus mariqueter, a habitat that provides a variety of living spaces and sufficient food sources for macrobenthic communities. Transect N4 presented various habitats such as dam, rock reef, gravel and mudflat, which contribute to the heterogeneity of substrates and provide a suitable environment and the preferred substrate for sessile or burrowing benthos, and for tide-dependent reptile benthic fauna (Zhong et al., 2020). Similarly, the species and functional diversity values for the macrobenthos in these two natural tidal flat areas were high. In contrast, due to the environmental modifications caused by ecological optimization engineering, communication with the natural water system were inadequate in the restoration area. In addition, with the inflow of fresh water and the water level rise, the salinity in the area decreased significantly, resulting in the growth of reeds, Scirpus mariqueter and submerged plants. In contrast, the changes in salinity in the natural tidal flats were relatively stable and had a limited effect on the original benthic communities. Previous studies have also found that salinity preference is a major functional feature that affects species distribution (van der Linden et al., 2012). In Sites S1, S1A and S3, river channels and different vegetation communities increased habitat heterogeneity, and the fresh water inflow was particularly beneficial to the survival of a number of species adapted to the low salinity environment, such as Stenothyra glabra, Elachisina sp., and Chinese white prawn, which dominated in these three restoration zones. These results are similar to those obtained in other habitats characterized by freshwater inflows (Yang et al., 2017; Lv et al., 2019). However, in Sites S2 and S2E, species recovery did not seem optimal. The replenishment rate of freshwater species was slower than the disappearance rate of the original benthos. In addition, the functional characteristics of the macrobenthos tended to be homogenized, which is also the result of the succession of macrobenthic communities caused by the environmental pressure under reclamation practices. Especially in Site S2, the development of the benthic fauna was inhibited in the bottom habitats, whose substrates were characterized by hypoxia because of the high density of reed and humus decomposition. Furthermore, the thick and hard rhizomes of reeds in the area do not represent a food source for benthic animals. Consequently, species diversity in Site S2 was significantly lower than that in the natural tidal flat, and both the functional diversity and dispersion also experienced a chain decline, indicating that the ecological function was significantly weakened. The utilization of environmental resources by the communities showing low functional differentiation decreased accordingly.
Functional redundancy reflects the replacement or absence of functional niches in a community after the disappearance of a species. This functional attribute is represented by FDQ/H', and the higher the ratio, the lower the functional redundancy. FDQ/H' was not significantly different between the restoration area and the natural tidal flat, but the ratio in the restoration area was higher; specifically, the highest value was reached in Site S2. This indicates that the negative effects of ecological optimization engineering on the succession of macrobenthic communities still exist. Even though the functional niche in the area was replaced by a number of freshwater species, the functional redundancy was reduced. If the decline of the existing macrobenthos persists in the region, it may negatively impact the stability of ecosystem functions. Environmental changes resulting from restoration projects alter the interactions between macrobenthic species in the original niche, which in turn may affect biodiversity in the restoration areas (Crain et al., 2004; Mor et al., 2018).
4.3
Comparison of ecological quality of different benthic habitats after reclamation
The funnel plot showed that the selected sampling points all fell within the 95% confidence interval, meaning that the classification of the macrobenthic species in the natural tidal flat and restoration areas remained unchanged. The values obtained for the M-AMBI indices showed that the ecological quality of natural tidal flat area was relatively high. However, the ABC curve and W value indicated that the disturbance degree of Transect N3 was low, while that of N4 was higher, defining this area as slightly disturbed. The high biodiversity of Transect N3 allowed the maintenance of a state of dynamic equilibrium during disturbance events. However, due to the scarcity of salt marsh vegetation, the barrier effect against tidal currents in Transect N4 was weakened and, due to the intense scouring action of seawater, the habitat of this area was defined as slightly disturbed. This result was consistent with previous studies (Liu et al., 2019). The AMBI values showed that all habitats were slightly disturbed. The M-AMBI results indicated that the ecological quality of Site S2 was poor, while that of S2E was good. Both restoration areas were planted with dense reed communities, which indicated that the ecological optimization project had caused some disturbance to the ecological quality of macrobenthic communities. In Site S2, it was observed that the developmental and ecological functions of macrobenthic communities were inhibited, leading to the degradation of the ecological quality. In the disturbed restoration areas, chironomid larvae (AMBI ecological group III) and Corophium volutator (AMBI ecological group III) presented adaptive advantages and could quickly establish population dominance in the disturbed habitats, benefiting from their small size, short life cycle and strong reproductive capacity. Nevertheless, the establishment of such dominant populations did not contribute satisfactorily to the restoration of the benthic community structure and ecological functions. In particular, the ecological quality in these disturbed areas was usually inferior, by the classification standards of ecological groups based on species. The ecological quality of Sites S1, S1A and S3 was higher in the restoration area, which may be due to the more stable function of macrobenthic communities and the stronger ability to withstand environmental changes.
Consequently, wetland managers should comprehensively analyze the changes and functional characteristics of the macrobenthos when defining ecological protection red line (nature reserve planning), and evaluate the ecological quality of wetland reserves, which will assist managers in formulating a more targeted implementation of ecological protection programs. By comparing the data before and after reclamation, it can be seen that the number of species, quantity, and diversity of the macrobenthos was shown to increase significantly. These results indicated that the ecological restoration of the macrobenthic community was fulfilled after the establishment of the ecological optimization project. In the present study, the species richness, density and biomass of the macrobenthos in the restoration area were still lower than those in the natural tidal flat, but species evenness, taxonomic characteristics, and community function of resource utilization in the restoration area were not different from those in the natural tidal flat. Differences in the composition and functional characteristics of the macrobenthic communities in restoration areas and natural tidal flats were closely related to complex habitat heterogeneity and salinity changes. In the restoration area, freshwater species compensated for the missing ecological niche of the original marine benthos, and the ecological functions in the area were thus restored to some extent. Nevertheless, the functional redundancy in the region was reduced, indicating that the effect of the functional replacement of this niche may not last long. In the ecological quality evaluation, the natural tidal flat area was also affected by tidal currents, which contributed to habitat instability. The ecological quality was relatively optimal in several restoration areas, but Site S2 and S2E were still moderately or slightly disturbed. Given the changes in ecological functions and ecological quality observed in the restoration areas, it is recommended that the administrative departments strengthen the management and protection of the restoration areas. In particular, measures should be taken to protect biodiversity in the restoration areas, such as harvesting reeds to reduce density, introducing fresh water species that can adapt to a low salinity environment, improving the macrobenthic habitat, and increasing its community functions and resistance to environmental disturbance, in order to accelerate the restoration and succession of the macrobenthic communities.
5.
Conclusions
This study compared data before and after reclamation, and found that the removal of S. alterniflora in the ecological optimization project was beneficial to the restoration of macrobenthic populations and community density, and resulted in good ecological benefits. Compared with the natural tidal-flat area, the extinction of the original macrobenthos in the restoration areas resulted in a significant decline in species richness, density, and biomass. These results indicated that the composition of macrobenthic communities in the natural tidal-flat area and the restoration area were clearly differentiated. The species biological trait of “salinity preference” affected the changes in the species composition and community structure of the macrobenthos in the study area. In the natural tidal-flat area, the biodiversity was in a dynamic equilibrium state, but Transect N3 was classified as undisturbed and Transect N4 as good quality. Although some restoration areas showed a downward trend in species richness, their classification and functional diversity were similar to those of the natural tidal flats, which indicated that the ecological functions of the two studied areas were similar; however, the functional distribution of specific species was significantly different. The reason for this may be that the different habitat heterogeneity present in the natural tidal flat had promoted a different succession of macrobenthos in the restoration area. The vegetation and freshwater environment in the restoration area provided a favorable living environment for freshwater species, which were also associated with species composition and, in part, with the restoration of ecological functions. However, the significant decline in species richness, density, biomass and biodiversity observed in Site S2 also indicated that the restoration effect was still not optimal, and that the speed with which species succession and niche compensation occurred was of great significance to the restoration of ecological functions. Although the ecological environment in part of the restoration areas was of good quality, the ecological health of Sites S2 and S2E was still critical, indicating that the resistance of macrobenthic communities to environmental changes needs to be strengthened. In addition to the disturbance caused by ecological optimization projects, high-density reed communities were also a main factor that affected the diversity and ecological quality of benthic communities. In order to improve the resistance of the benthic fauna to environmental disturbance, management policies should focus on protecting the biological diversity and the functional stability of the ecosystem in the restoration area. For example, reeds can be harvested and freshwater species can be introduced to create favorable conditions for the recovery of benthic communities.
Acknowledgements:
We thank Xuemei Han of Nanyang Normal University and Wanxiang Jiang of Zaozhuang University for their guidance and assistance in the calculation of the functional diversity index. We thank Yuwei Huang of South China Normal University for translating and proofreading the list of macrozoobenthic species in the Changjiang River Estuary. We thank International Science Editing (http://www.internationalscienceediting.com) for editing this manuscript.
Adler P B, Bradford J B. 2002. Compensation: an alternative method for analyzing diversity-productivity experiments. Oikos, 96(3): 411–420. doi: 10.1034/j.1600-0706.2002.960303.x
Barbier E B, Hacker S D, Kennedy C, et al. 2011. The value of estuarine and coastal ecosystem services. Ecological Monographs, 81(2): 169–193. doi: 10.1890/10-1510.1
Bertness M D, Ewanchuk P J, Silliman B R. 2002. Anthropogenic modification of New England salt marsh landscapes. Proceedings of the National Academy of Sciences of the United States of America, 99(3): 1395–1398. doi: 10.1073/pnas.022447299
Borja A, Dauer D M, Díaz R, et al. 2008. Assessing estuarine benthic quality conditions in Chesapeake Bay: A comparison of three indices. Ecological Indicators, 8(4): 395–403. doi: 10.1016/j.ecolind.2007.05.003
Borja Á, Dauer D M, Grémare A. 2012. The importance of setting targets and reference conditions in assessing marine ecosystem quality. Ecological Indicators, 12(1): 1–7. doi: 10.1016/j.ecolind.2011.06.018
Borja A, Franco J, Pérez V. 2000. A marine Biotic Index to establish the ecological quality of soft-bottom benthos within European estuarine and coastal environments. Marine Pollution Bulletin, 40(12): 1100–1114. doi: 10.1016/s0025-326x(00)00061-8
Borja Á, Marín S L, Muxika I, et al. 2015. Is there a possibility of ranking benthic quality assessment indices to select the most responsive to different human pressures?. Marine Pollution Bulletin, 97(1–2): 85–94,
Chen Zhongbing, Guo Li, Jin Binsong, et al. 2009. Effect of the exotic plant Spartina alterniflora on macrobenthos communities in salt marshes of the Yangtze River Estuary, China. Estuarine, Coastal and Shelf Science, 82(2): 265–272,
Chen Jing, Jiang Wanxiang, He Shishui, et al. 2018. Study of macroinvertebrate species and functional diversity in the New Xue River, Shandong Province, China. Acta Ecologica Sinica, 38(9): 3328–3336
Christianen M J A, Middelburg J J, Holthuijsen S J, et al. 2017. Benthic primary producers are key to sustain the Wadden Sea food web: stable carbon isotope analysis at landscape scale. Ecology, 98(6): 1498–1512. doi: 10.1002/ecy.1837
Crain C M, Silliman BR, Bertness S L, et al. 2004. Physical and biotic drivers of plant distribution across estuarine salinity gradients. Ecology, 85(9): 2539–2549. doi: 10.1890/03-0745
Forde James, O’Beirn Francis X, O’Carroll Jack P J, et al. 2015. Impact of intertidal oyster trestle cultivation on the ecological status of benthic habitats. Marine Pollution Bulletin, 95(1): 223–233. doi: 10.1016/j.marpolbul.2015.04.013
Gao Yu, Lin Guanghui. 2018. Algal diversity and their importance in ecological processes in typical mangrove ecosystems. Biodiversity Science, 26(11): 1223–1235. doi: 10.17520/biods.2018080
Gao Z G, Zhang L Q. 2006. Multi-seasonal spectral characteristics analysis of coastal salt marsh vegetation in Shanghai, China. Estuarine, Coastal and Shelf Science, 69(1–2): 217–224,
Han Taotao, Tang Xuan, Ren Hai, et al. 2021. Community/ecosystem functional diversity: measurements and development. Acta Ecologica Sinica, 41(8): 3286–3295
Herman P M J, Middelburg J J, Van De Koppel J, et al. 1999. Ecology of estuarine macrobenthos. In: Nedwell D B, Raffaelli D G, eds. Advances in Ecological Research. Academic Press, 29: 195–240
Huang Yinying, Li Yiming, Chen Qiang, et al. 2021. Effects of reclamation methods and habitats on macrobenthic communities and ecological health in estuarine coastal wetlands. Marine Pollution Bulletin, 168: 112420. doi: 10.1016/j.marpolbul.2021.112420
Korb S K, Bolshakov L V, Fric Z F, et al. 2016. Cluster biodiversity as a multidimensional structure evolution strategy: checkerspot butterflies of the group Euphydryas aurinia (Rottemburg, 1775) (Lepidoptera: Nymphalidae). Systematic Entomology, 41(2): 441–457. doi: 10.1111/syen.12167
Laursen A E, Seitzinger S P, Dekorsey R, et al. 2002. Multiple stressors in an estuarine system: Effects of nutrients, trace elements, and trophic complexity on benthic photosynthesis and respiration. Estuaries, 25(1): 57–69. doi: 10.1007/BF02696049
Leung J Y S. 2015. Habitat heterogeneity affects ecological functions of macrobenthic communities in a mangrove: Implication for the impact of restoration and afforestation. Global Ecology and Conservation, 4: 423–433. doi: 10.1016/j.gecco.2015.08.005
Levins R. 1970. An introduction to mathematical ecology. Evolution, 24(2): 482
Li Xiaoxiao, Yang Wei, Sun Tao, et al. 2019. Framework of multidimensional macrobenthos biodiversity to evaluate ecological restoration in wetlands. Environmental Research Letters, 14(5): 054003. doi: 10.1088/1748-9326/ab142c
Liu Zhiquan, Chen Minghai, Li Yiming, et al. 2018a. Different effects of reclamation methods on macrobenthos community structure in the Yangtze Estuary, China. Marine Pollution Bulletin, 127: 429–436. doi: 10.1016/j.marpolbul.2017.12.038
Liu Zhiquan, Fan Bin, Huang Youhui, et al. 2019. Assessing the ecological health of the Chongming Dongtan Nature Reserve, China, using different benthic biotic indices. Marine Pollution Bulletin, 146: 76–84. doi: 10.1016/j.marpolbul.2019.06.006
Liu Zhiquan, Yu Ping, Chen Minghai, et al. 2018b. Macrobenthic community characteristics and ecological health of a constructed intertidal oyster reef in the Yangtze Estuary, China. Marine Pollution Bulletin, 135: 95–104. doi: 10.1016/j.marpolbul.2018.07.019
Luo Xianxiang, Sun Kaijing, Yang Jianqiang, et al. 2016. A comparison of the applicability of the Shannon-Wiener index, AMBI and M-AMBI indices for assessing benthic habitat health in the Huanghe (Yellow River) Estuary and adjacent areas. Acta Oceanologica Sinica, 35(6): 50–58. doi: 10.1007/s13131-016-0842-9
Lv Weiwei, Huang Youhui, Liu Zhiquan, et al. 2016a. Application of macrobenthic diversity to estimate ecological health of artificial oyster reef in Yangtze Estuary, China. Marine Pollution Bulletin, 103(1–2): 137–143,
Lv Weiwei, Liu Zhiquan, Yang Yang, et al. 2016b. Loss and self-restoration of macrobenthic diversity in reclamation habitats of estuarine islands in Yangtze Estuary, China. Marine Pollution Bulletin, 103(1–2): 128–136,
Lv Weiwei, Zhou Wenzong, Zhao Yunlong. 2019. Effect of freshwater inflow on self-restoration of macrobenthic diversity in seaward intertidal wetlands influenced by reclamation projects in the Yangtze Estuary, China. Marine Pollution Bulletin, 138: 177–186. doi: 10.1016/j.marpolbul.2018.11.044
Ma Qiang, Wu Wei, Tang Chendong, et al. 2017. Effects of habitat restoration on the diversity of bird and marcobenthos in the Chongming Dongtan wetland. Journal of Nanjing Forestry University (Natural Sciences Edition), 41(1): 9–14
Margalef R. 1969. Diversity and stability: a practical proposal and a model of interdependence. Brookhaven Symposia in Biology, 22: 25–37
Martinez-Haro M, Beiras R, Bellas J, et al. 2015. A review on the ecological quality status assessment in aquatic systems using community based indicators and ecotoxicological tools: what might be the added value of their combination?. Ecological Indicators, 48: 8–16. doi: 10.1016/j.ecolind.2014.07.024
Mascaro J, Hughes R F, Schnitzer S A. 2012. Novel forests maintain ecosystem processes after the decline of native tree species. Ecological Monographs, 82(2): 221–228. doi: 10.1890/11-1014.1
Mor J R, Ruhí A, Tornés E, et al. 2018. Dam regulation and riverine food-web structure in a Mediterranean river. Science of the Total Environment, 625: 301–310. doi: 10.1016/j.scitotenv.2017.12.296
Muxika I, Borja Á, Bald J. 2007. Using historical data, expert judgement and multivariate analysis in assessing reference conditions and benthic ecological status, according to the European Water Framework Directive. Marine Pollution Bulletin, 55(1–6): 16–29,
Nie Minghua, Yan Caixia, Dong Wenbo, et al. 2015. Occurrence, distribution and risk assessment of estrogens in surface water, suspended particulate matter, and sediments of the Yangtze Estuary. Chemosphere, 127: 109–116. doi: 10.1016/j.chemosphere.2015.01.021
Nishijima W, Nakano Y, Nakai S, et al. 2014. Macrobenthic succession and characteristics of a man-made intertidal sandflat constructed in the diversion channel of the Ohta River Estuary. Marine Pollution Bulletin, 82(1–2): 101–108,
Nunes M, Coelho J P, Cardoso P G, et al. 2008. The macrobenthic community along a mercury contamination in a temperate estuarine system (Ria de Aveiro, Portugal). Science of The Total Environment, 405(1–3): 186–194,
Petchey O L. 2003. Integrating methods that investigate how complementarity influences ecosystem functioning. Oikos, 101(2): 323–330. doi: 10.1034/j.1600-0706.2003.11828.x
Petchey O L, Gaston K J. 2002. Functional diversity (FD), species richness and community composition. Ecology Letters, 5(3): 402–411. doi: 10.1046/j.1461-0248.2002.00339.x
Petchey O L, Gaston K J. 2006. Functional diversity: back to basics and looking forward. Ecology Letters, 9(6): 741–758. doi: 10.1111/j.1461-0248.2006.00924.x
Peterson C H, Lipcius R N. 2003. Conceptual progress towards predicting quantitative ecosystem benefits of ecological restorations. Marine Ecology Progress Series, 264: 297–307. doi: 10.3354/meps264297
Piló D, Ben-Hamadou R, Pereira F, et al. 2016. How functional traits of estuarine macrobenthic assemblages respond to metal contamination?. Ecological Indicators, 71: 645–659. doi: 10.1016/j.ecolind.2016.07.019
Poos M S, Walker S C, Jackson D A. 2009. Functional-diversity indices can be driven by methodological choices and species richness. Ecology, 90(2): 341–347. doi: 10.1890/08-1638.1
Ren Zhonghua, Li Fan, Wei Jiali, et al. 2016. Community characteristics of macrobenthos in the Huanghe (Yellow River) Estuary during water and sediment discharge regulation. Acta Oceanologica Sinica, 35(8): 74–81. doi: 10.1007/s13131-016-0881-2
Rosenfeld J S. 2002. Functional redundancy in ecology and conservation. Oikos, 98(1): 156–162. doi: 10.1034/j.1600-0706.2002.980116.x
Sakamaki T, Richardson J S. 2009. Dietary responses of tidal flat macrobenthos to reduction of benthic microalgae: a test for potential use of allochthonous organic matter. Marine Ecology Progress Series, 386: 107–113. doi: 10.3354/meps08094
Salas F, Marcos C, Neto J M, et al. 2006. User-friendly guide for using benthic ecological indicators in coastal and marine quality assessment. Ocean & Coastal Management, 49(5–6): 308–331,
Schmera D, Erős T, Podani J. 2009. A measure for assessing functional diversity in ecological communities. Aquatic Ecology, 43(1): 157–167. doi: 10.1007/s10452-007-9152-9
Seitzinger S. 2008. Out of reach. Nature, 452(7184): 162–163. doi: 10.1038/452162a
Shannon C E, Weaver W. 1962. The Mathematical Theory of Communication. Urbana: University of Illinois Press
Tang Chendong. 2016. Ecological control of Spartina alternilfora and improvement of birds habitats in Chongming Dongtan Wetland, Shanghai. Wetland Science & Management, 12(3): 4–8
Tian Bo, Zhou Yunxuan, Zhang Liquan, et al. 2008. Analyzing the habitat suitability for migratory birds at the Chongming Dongtan Nature Reserve in Shanghai, China. Estuarine, Coastal and Shelf Science, 80(2): 296–302,
Tilman D, Reich P B, Knops J, et al. 2001. Diversity and productivity in a long-term grassland experiment. Science, 294(5543): 843–845. doi: 10.1126/science.1060391
Usseglio-Polatera P, Bournaud M, Richoux P, et al. 2000. Biological and ecological traits of benthic freshwater macroinvertebrates: relationships and definition of groups with similar traits. Freshwater Biology, 43(2): 175–205. doi: 10.1046/j.1365-2427.2000.00535.x
van der Linden P, Patrício J, Marchini A, et al. 2012. A biological trait approach to assess the functional composition of subtidal benthic communities in an estuarine ecosystem. Ecological Indicators, 20: 121–133. doi: 10.1016/j.ecolind.2012.02.004
Voß K, Schäfer R B. 2017. Taxonomic and functional diversity of stream invertebrates along an environmental stress gradient. Ecological Indicators, 81: 235–242. doi: 10.1016/j.ecolind.2017.05.072
Wang Ruizhao, Yuan Lin, Zhang Liquan. 2010. Impacts of Spartina alterniflora invasion on the benthic communities of salt marshes in the Yangtze Estuary, China. Ecological Engineering, 36(6): 799–806. doi: 10.1016/j.ecoleng.2010.02.005
Wang Xuejing, Zhang Yan, Luo Manhua, et al. 2021. Radium and nitrogen isotopes tracing fluxes and sources of submarine groundwater discharge driven nitrate in an urbanized coastal area. Science of the Total Environment, 763: 144616. doi: 10.1016/j.scitotenv.2020.144616
Warwick R M. 1986. A new method for detecting pollution effects on marine macrobenthic communities. Marine Biology, 92(4): 557–562. doi: 10.1007/BF00392515
Warwick R M, Clarke K R. 1994. Relearning the ABC: taxonomic changes and abundance/biomass relationships in disturbed benthic communities. Marine Biology, 118(4): 739–744. doi: 10.1007/BF00347523
Witman J D, Cusson M, Archambault P, et al. 2008. The relation between productivity and species diversity in temperate-arctic marine ecosystems. Ecology, 89(S11): S66–S80. doi: 10.1890/07-1201.1
Wong M C, Dowd M. 2015. Patterns in taxonomic and functional diversity of macrobenthic invertebrates across seagrass habitats: a case study in Atlantic Canada. Estuaries and Coasts, 38(6): 2323–2336. doi: 10.1007/s12237-015-9967-x
Yang Wei, Li Ming, Sun Tao, et al. 2017. The joint effect of tidal barrier construction and freshwater releases on the macrobenthos community in the northern Yellow River Delta (China). Ocean & Coastal Management, 136: 83–94. doi: 10.1016/j.ocecoaman.2016.11.024
Zhong Xin, Qiu Baochao, Liu Xiaoshou. 2020. Functional diversity patterns of macrofauna in the adjacent waters of the Yangtze River Estuary. Marine Pollution Bulletin, 154: 111032. doi: 10.1016/j.marpolbul.2020.111032
Zhou Zhengquan, Li Xiaojing, Chen Linlin, et al. 2018. Macrobenthic assemblage characteristics under stressed waters and ecological health assessment using AMBI and M-AMBI: a case study at the Xin’an River Estuary, Yantai, China. Acta Oceanologica Sinica, 37(5): 77–86. doi: 10.1007/s13131-018-1180-x
Zhu Xiaofen, Chen Bin, Yu Weiwei, et al. 2018. Discussion on taxonomic diversity indices and taxonomic sufficiency for macrobenthos in Xiamen Bay. Acta Ecologica Sinica, 38(15): 5554–5565
Changqing Liu, Hai Cheng, Yixin Wu, et al. Evaluation of soil fauna biodiversity in restored farmland for protection of wetland ecology by planting different crops. Ocean & Coastal Management, 2024, 247: 106945. doi:10.1016/j.ocecoaman.2023.106945
2.
Steffany Katherine Baudisch, Kalina Manabe Brauko, Paulo da Cunha Lana. Determination of organic contamination levels by the ABC Method (Abundance/Biomass Curves) in intertidal estuarine flats using hierarchical design. Ocean and Coastal Research, 2024, 72(suppl 1) doi:10.1590/2675-2824072.23143
3.
Satyajit Paul, Swades Pal. Impact of wetland health and provisioning services on the livelihood of the fishing community. Environmental Science and Pollution Research, 2024, 31(39): 52410. doi:10.1007/s11356-024-34573-3
4.
Xinglin Du, Yizhou Huang, Yucong Ye, et al. Effects of variations in hydrological connectivity on the macrobenthic community structure in reclaimed wetlands. Science of The Total Environment, 2024, 954: 176111. doi:10.1016/j.scitotenv.2024.176111
5.
Cheng Wang, Shaoxia Xia, Houlang Duan, et al. Study on the impact of reclamation and development of Yancheng coastal wetlands on the spatio-temporal evolution of wintering Anatidae habitat. Ecological Indicators, 2024, 160: 111892. doi:10.1016/j.ecolind.2024.111892
6.
Yinan Zhang, Hang Wan, Yunlong Zhao, et al. Macrobenthic functional group analysis of ecological health of the intertidal artificial oyster reefs in the Yangtze Estuary, China. Frontiers in Marine Science, 2023, 9 doi:10.3389/fmars.2022.1059353
7.
Xue Mo, Jingqiu Chen, Yueyan Pan, et al. Impact of applying imazapyr on the control of Spartina alterniflora and its eco-environments in the Yellow River Delta, China. Watershed Ecology and the Environment, 2022, 4: 211. doi:10.1016/j.wsee.2022.11.002
Table
2.
The biodiversity index of macrobenthos in the Chongming Dongtan wetland, Shanghai from 2019 to 2020
Transect N3
Transect N4
Site S1
Site S1A
Site S2
Site S2E
Site S3
DO concentration/(mg·L−1)
7.95±2.72
7.89±4.54
8.40±3.50
10.09±4.92
11.45±3.84
8.63 ± 2.69
10.49 ± 2.61
pH
8.14±0.47
7.88±0.52
8.02±0.49
8.32±0.29
8.36±0.49
7.96±0.75
8.09±0.50
Water temperature/°C
20.72±10.11
20.30±10.04
19.73±7.77
19.82±8.41
20.49±8.13
19.71±8.38
20.52±8.64
Salinity
8.33±1.91b
8.77±2.27b
3.52±2.65a
3.46±2.67a
1.37±2.07a
3.11±1.97a
4.37±1.96a
d
2.18±0.59c
1.85±0.59bc
0.94±0.42ab
1.67±0.87bc
0.19±0.26a
1.44±0.60bc
1.23±0.74b
J'
0.62±0.15ab
0.65±0.14b
0.70±0.15b
0.82±0.10b
0.18±0.29a
0.60±0.19ab
0.77±0.15b
H'
2.36±0.65b
2.31±0.59b
1.62±0.39b
2.54±0.81b
0.31±0.59a
1.86±0.72b
2.01±1.00b
Δ+
88.33±3.73b
83.43±14.02b
84.74±9.01b
82.47±9.55b
48.26±51.67a
85.24±2.70b
90.13±6.84b
$\varLambda $+
437.13±108.99b
362.08±65.10ab
331.40±166.21ab
533.50±414.53b
51.12±121.58a
521.72±113.81b
332.89±234.43ab
FDQ
42.56±3.20b
44.97±4.10b
37.81±7.68b
41.88±6.15b
11.99±13.81a
38.92±6.67b
39.23±11.46b
FDis
6.42±0.25b
6.60±0.33b
6.01±0.69b
6.39±0.49b
2.20±2.47a
6.15±0.55b
6.09±1.09b
FDQ/H'
19.38±6.34
20.86±7.37
24.49±7.44
17.89±5.95
39.50±53.30
25.03±12.74
24.54±14.46
Note: d is the Margalef species richness index; J', Pielou species evenness index; H', Shannon-Wiener index; Δ+, average taxonomic distinctness; $\varLambda $+, variation in taxonomic distinctness; FDQ, functional diversity index; FDis, functional dispersion; and FDQ/H', functional redundancy. Different letters indicate significant differences among sites.
Figure 1. Schematic diagram of macrobenthic sampling in Chongming Dongtan wetland from 2019 to 2020. In b, the dark grey area represents the semi-closed reclamation restoration area, and the light grey area represents the natural tidal-flat area. The dots represent the sampling sites; the dash lines represent the Transects N3 and N4.
Figure 2. The species composition (a), number of species (b), density (c) and biomass (d) of macrobenthos in Chongming Dongtan wetland from 2019 to 2020. N3 and N4 are sampling transects; S1, S1A, S2, S2E and S3 are sampling sites. Different letters above the bars indicate significant differences among sites.
Figure 3. The correlation matrix of the macrobenthic diversity index in Chongming Dongtan wetland from 2019 to 2020. The d is the Margalef species richness index; J′, Pielou species evenness index; H', Shannon-Wiener index; Δ+, average taxonomic distinctness; Λ+, variation in taxonomic distinctness; FDQ, functional diversity index; FDis, functional dispersion; and FDQ/H', functional redundancy. Species diversity includes d, J' and H'; taxonomic diversity, Δ+ and Λ+; functional diversity, FDQ and FDis; Functional redundancy, FD/H'.
Figure 4. Clustering and non-metric multidimensional scaling (NMDS) analysis of macrobenthic communities in Chongming Dongtan wetland from 2019 to 2020.
Figure 5. Abundance-biomass comparison curves of 7 habitats in Chongming Dongtan Wetland from 2019 to 2020 (green triangle is abundance; blue triangle, biomass).




 DownLoad:
DownLoad:
 DownLoad:
DownLoad:









 DownLoad:
DownLoad: